Chap-9-limmunité-adaptative-le-prolongement-de

Chap. 9 L’immunité adaptative, prolongement de l’immunité innée
En plus de l’immunité innée immédiatement efficace contre de nombreux agresseurs, les
vertébrés possèdent une immunité adaptative (ou acquise) plus efficace mais qui ne devient
performante qu’après un premier contact avec l’antigène.
9.1 L’immunité adaptative, une immunité spécifique
TD 9.1 : Act 1p310
Correction :
Doc. 1 : Dans l’expérience 1, les cobayes ayant reçu l’injection d’un « vaccin approprié »
(anatoxine tétanique) sont protégés contre le tétanos mais pas contre la diphtérie ni contre
toute autre toxine microbienne. On peut donc dire que la protection assurée par l’injection
d’anatoxine tétanique est spécifique.
Doc. 2 : Dans les expériences 2 et 3, le cobaye A survit à une injection de l’agent pathogène
contre lequel il a été immunisé. L’immunisation lui a donc conféré une immunité contre cet
agent pathogène. Une expérimentation comparable à celle présentée dans Partie 4. Corps
196 humain et santé
le doc. 1 montrerait que, là encore, cette immunité est spécifique.
Dans les deux cas, on essaie ensuite de transférer de manière passive, par injection de sérum
ou de cellules, l’immunité d’un animal immunisé (cobaye B ou C) à un animal non immunisé
(cobaye D ou E).
Dans l’expérience 2, la protection est obtenue par transfert de sérum ; on dit que c’est une
défense à médiation humorale (en vieux français, le mot « humeurs » désignait
l’ensemble des liquides de l’organisme).
Dans l’expérience 3, la protection est obtenue par transfert de cellules ; on dit que
c’est une défense à médiation cellulaire.
Doc. 3 et 4 : Les lymphocytes sont les cellules immunitaires qui assurent les défenses
spécifiques et sont donc le support de l’immunité adaptative. Pour aboutir à l’élimination d’un
agent pathogène, il y a toujours nécessité d’une coopération entre différentes catégories de
lymphocytes.
En effet, c’est dans le cas où les trois types de lymphocytes sont présents que l’efficacité
immunitaire est maximale. Elle est moindre en l’absence des seuls T CD8 ou B.
En revanche, l’absence simultanée de deux types de lymphocytes (et a fortiori des trois
types) ne permet pas une défense immunitaire efficace contre le virus grippal.
Cette première approche pose le problème du rôle exact de chaque catégorie lymphocytaire,
ainsi que celui du mécanisme de coopération entre les différents types cellulaires.
Bilan :
L’immunité adaptative est une immunité spécifique car les
mécanismes de défense mis en jeu sont dirigés contre un seul
type d’agent pathogène qui est reconnu de manière spécifique.
La protection de l’organisme contre cet agent pathogène peut
être assurée soit par des substances solubles dans le plasma
sanguin (immunité à médiation humorale), soit directement par
certaines catégories de lymphocytes (immunité à médiation
cellulaire).

Les lymphocytes sont le support de cette immunité adaptative
mais l’élimination d’un agent pathogène nécessite toujours la
coopération entre différentes catégories de lymphocytes.
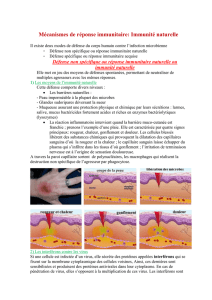
9.2 La réponse adaptative à médiation humorale au virus de la grippe
TD 9.2: Infection par le virus de la grippe
Comme tous les virus, le virus de la grippe doit parasiter certaines de nos cellules : ici, les
cellules épithéliales des voies respiratoires.
Pour reconnaître et pénétrer dans nos cellules, les virus disposent à leur surface de protéines
spécifiques (Ha : hémaglutinine, Na : neuramidase) qui seront reconnus par notre système
immunitaire et constitueront pour nous des antigènes.
Avant de parasiter une cellule, le virus se déplace dans le sang ou la lymphe.
Il est alors accessible aux lymphocytes B qui sont les cellules de la médiation humorale. Les
Lb possèdent à leur surface des anticorps membranaires.
TP1 9.2 Visualisation avec rastop d'une immunoglobuline globale +
paratope et épitope
Un Ac (ou immunoglobuline) est une protéine en forme de Y constituée de 4 chaines d’acides
aminés identiques 2 à 2 : 2 chaines lourdes (H) et 2 chaines légères (L). Chaque chaine
possède une partie constante et une partie variable d’un Ac à un autre : c’est cette partie qui
est responsable de la spécificité.
Il y a donc 2 sites de fixation à l’Ag (aux extrémités des branches) et un site de fixation aux
phagocytes (base du Y).
TP2 9.2 spécificité Ac avec rastop
TP3 9.2 Ouchterlony
Chaque Lb ne porte qu’un seul
type d’Ac et ne peut donc
détecter qu’un seul Ag, mais
nous disposons dans notre
organismes de millions de
clones de Lb différents.

Suite à la reconnaissance par un Lb de son Ag, le Lb est sélectionné (phase de sélection
clonale).
En présence de la molécule interleukine 2 (Il2), les Lb sélectionnés vont se multiplier par
mitose (phase d’amplification clonale) puis se différencier pour certains en Lb mémoire à
longue durée de vie et pour d’autres en plasmocytes sécréteurs d’Ac circulants (phase de
différenciation clonale).
Schéma 3p315.
Les Ac libérés dans le plasma reconnaissent le même Ag que les Ac membranaires du Lb à
l’origine du plasmocyte.
Schéma 4p317
La partie constante des Ac est reconnue par des récepteurs spécifiques des phagocytes qui
vont alors ingérer et détruire le complexe immun (donc l’Ag)
9.3 La réponse immunitaire à médiation cellulaire.
Quand l’Ag (virus de la grippe) pénètre dans une nos cellules (voies respiratoires), il ne peut
plus être combattu par les Ac produits par les LB.
Au cours de la réaction immunitaire innée, les cellules dendritiques ont phagocytés et digérés
l’Ag. Elles en présentent maintenant des parties dans leur récepteur CMH et sont devenues
des cellules présentatrices d’antigènes : CPA.
Schéma 2p319
Les LTCD8 disposent de récepteurs spécifiques capables de reconnaître un Ag présenté dans le
CMH par une CPA.
Il existe des millions de LTCD8 différents par leur récepteur, chaque LTCD8 ne reconnaissant
qu’un Ag.
La reconnaissance entre un Ag et un LTCD8 spécifique provoque la phase de sélection
clonale.
En présence d’interleukine 2 (IL2), le LTCD8 sélectionné se multiplie par mitose : c’est la
phase d’amplification clonale.

Ensuite les LTCD8 se différencient:
- soit en LTcytotoxiques (LTc) qui quittent les ganglions lymphatiques
- soit en LTc mémoire à longue durée de vie
C’est la phase de différenciation.
Quand les LTc rencontrent une cellule infectée par le même Ag, ils la reconnaissent car elle
présente cet Ag sur sa membrane (schéma 1p318).
Ils la détruisent alors :
- soit par cytolyse (ils font des trous dans la membrane)
- soit par apoptose (ils forcent sa destruction).
9.4 Les LTCD4+
9.4.1 Au centre des réactions adaptatives.
Voir schémas bilans postés
Les LTCD4, tout comme les LTCD8 possèdent des récepteurs T spécifiques.
En présence d’une CPA présentant le bon Ag, ils sont sélectionnés, se multiplient et se
différencient (sans avoir besoin d’IL2).
Certains donnent des LT4 mémoire à longue durée de vie, d’autres donnent des LThelpers (LTh)
qui sécrètent IL2, indispensable au LB et LTCD8.
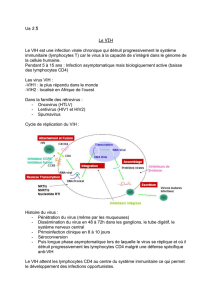
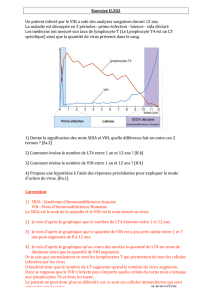
9.4.2 Cible du virus du SIDA
Entrée du VIH dans un LT cd4

L'infection virale en 3 étapes
La primo-infection
Il y a transmission du V.I.H selon 3 modes de contaminations: par voie sexuelle (70
à 80%) homo et hétérosexuelle parce que le virus est contenu dans les sécrétions génitales;
par voie sanguine (échange de seringues, transfusion, par piqûre accidentelle; de la mère
à l'enfant (contamination in-utéro pendant la grossesse à travers le placenta, pendant
l'allaitement)
Après la contamination, le virus se propage dans tout l'organisme via la circulation
sanguine et lymphatique. Il rentre en contact avec de nombreuses cellules dont les
lymphocytes B, T et les macrophages. Le virus comme la plupart des virus est un parasite
obligatoire des cellules dont il se sert afin de se reproduire. C'est un virus de l'ordre de
quelques dizaines à quelques centaines de nanomètres (si l'on voulait comparer la taille du
VIH à la taille d'une cellule, la taille du VIH correspondrait à la taille d'une prise USB
comparée à une feuille A4).
Il est constitué d'un génome deux simples chaînes d'ARN protégées par deux coques
protéiques ou capsides. Le tout contenu dans une enveloppe externe en bicouche lipidique
dans laquelle sont incrustées des glycoprotéines.
Le VIH ne va infecter que les lymphocytes T4 et les macrophages parce que ce sont
les seuls qui possèdent des récepteurs CD4. Seule la protéine CD4 est reconnue par la
glycoprotéine 120 du virus. Seuls les LT4 et les macrophages porteurs de CD4 sont les
cellules cibles du VIH.
Les nouveaux virus libérés dans le milieu intérieur peuvent à leur tour infecter
d’autres cellules cibles. La charge virale augmente expliquant ainsi les symptômes de la
primo-infection. Les symptômes peuvent apparaître de 2 à 6 semaines après la contamination
et durent environ 2 semaines. Cette infection conduit à la baisse du nombre de LT4 et
déclenche du même coup une intense réaction immunitaire de la part de l’hôte avec
l’apparition dans le sérum de l’organisme infecté de protéines spécifiques appelées
immunoglobuline (Ig) correspondant à des anticorps spécifiques anti-GP120. Le nombre de
lymphocytes cytotoxiques (LTC ou L.killers) à récepteurs CD8 va augmenter. Ces LT8 vont
détruire les cellules infectées par le VIH dont les LT4. La prolifération du VIH est
momentanément jugulée et on assiste à une augmentation du nombre de LT4. La présence
d’anticorps anti VIH et de LT8 signe l’immunité acquise et la fin de la primo-infection.
 6
7
6
7
1
/
7
100%