Neuroanatomie : guide de l`examen- sujets faits

Neuroanatomie : guide de l’examen- sujets faits
1. Méninges :
Énumérez dans l’ordre les feuillets et espaces méningés et les insertions des deux principales
expansions de dure-mère.
Dans l’ordre le plus à l’extérieur juste en dedans de l’os on trouve la pachyméninge (c'est-à-
dire la duremère composée d’une couche externe et d’une couche interne) qui est adhérente à
l’os au niveau du crâne mais pas dans les vertèbres, puis beaucoup plus fine et près de
l’encéphale on trouve la leptoméninge constituée de l’arachnoïde (qui tapisse la face interne
de la duremère) et la pie mère qui adhère totalement à la surface du cerveau (elle est constitué
de l’intima pia et de la couche épipiale).
Il existe 3 espaces méningés : tout d’abord l’espace extra dural se trouveant entre la duremère
et l’os, puis on trouve l’espace subdural entre la duremère et l’arachnoïde et enfin l’espace
subarachnoïdien contenant le LCR entre l’arachnoïde et la pie mère. Les deux premiers sont
virtuels, le troisième est développé et forme des citernes.
Pour résumer on trouve de l’os à l’encéphale : espace extra duralduremèreespace
subdural arachnoïde espace sous arachnoïdien pie-mère.
La duremère est épaisse et émet des expansions qui divisent le volume intracrânien : la faux
du cerveau et la tente du cervelet. La faux du cerveau est une cloison médio sagittale,
falciforme, elle présente 2 faces latérales en rapport avec les faces internes des hémisphères
cérébraux. Elle présente une base qui s’insère sur la tente du cervelet ; un bord supérieur qui
s’insère en avant sur le foramen caecum, puis d’avant en arrière sur les deux lèvres de la
gouttière du sinus sagittal supérieur à la face endocrânienne de la voûte ; et un bord inférieur
qui s’insère en avant sur le processus crista Galli de l’os ethmoïde, puis d’avant en arrière ce
bord est libre. La tente du cervelet est une cloison transversale séparant la cavité crânienne en
étage sus et sous tentoriel ; son bord antérieur est libre et s’insère en avant sur le processus
clinoïde antérieur ; son bord postérieur s’insère d’avant en arrière sur le processus clinoïde
postérieur, le bord supérieur du rocher et la gouttière du sinus transverse.
2. Anatomie descriptive de la configuration externe de la moelle spinale : sillons cordons
racines.
La moelle spinale est un long cordon cylindrique aplati d’avant en arrière. Elle a pour limite
supérieure l’émergence de la première racine cervicale et pour limite inférieure la vertèbre L1
ou L2. Elle présente 2 intumescences (cervicale et lombale), entre elle la moelle thoracique,
en bas se trouve l’épicone, le cône médullaire et le filum terminal.
Six sillons longitudinaux subdivisent la moelle en six funiculus. On décrit la fissure médiane
ventrale qui est une large dépression sur la ligne médiane ventrale, le sillon médian dorsal qui
est peu marqué et des sillons latéraux (ventro latéral et dorso latéral) qui correspondent à la
ligne d’émergence des radicelles dont la réunion forme les racines ventrales et dorsales. La
face dorsale de la moelle cervicale est marquée par deux sillons intermédiaires dorsaux situés
entre le sillon médian dorsal en dedans et les sillons latéraux dorsaux en dehors.
Les racines ventrales formées par l’ensemble des radicelles émergeant du sillon ventrolatéral
sont motrices. Les racines dorsales pénetrent dans le sillon dorso-latéral, elles véhiculent les
informations sensitives, le ganglion spinal des racines dorsales contient le corps cellulaires
des neurones sensitifs. De part et d’autres de la moelle, les racines ventrales et dorsales se
rejoignent et forme le nerf rachidien qui émergent par les trous de conjugaison.
Les cordons sont des colonnes de substance blanche formées par les axones myélinisées des
voies descendantes, montantes et d’association. On décrit un cordon ou funiculus ventral entre
la fissure médiane ventrale et le sillon ventral latéral qui véhicule les voies descendantes

motrices et de la sensibilité thermo-algique, le cordon latéral entre les sillons ventro latéral et
dorso latéral qui contient les voies de la motricité et de la sensibilité thermo algique et le
cordon dorsal entre le sillon médian dorsal et les sillons dorso latéraux qui véhiculent les
voies ascendantes de la sensibilité épicritique (ou lemniscale), au dessus du segment
médullaire D2, ils se divisent en deux faisceaux séparés par les sillons paramédians dorsaux.
Les deux funiculus ventraux sont réunis par la commissure blanche ventrale en avant du canal
central.
3. Anatomie descriptive de la configuration interne de la moelle spinale.
La moelle spinale est la partie inférieure du névraxe, elle est située dans le rachis. Elle a gardé
un aspect primitif : elle a un aspect segmenté, la substance grise est centrale, elle entoure le
canal central alors que la substance blanche est situé en périphérie.
La substance grise est formée par les corps cellulaires des neurones, elle forme des centres
nerveux qui relaient l’information. La substance blanche est constituée de fibres nerveuses,
dont la myéline donne la coloration blanche.
Les cornes sont des colonnes de substance grise. On décrit la corne dorsale qui est réceptrice,
organisée en 6 lames numérotées de I à VI de la périphérie vers le centre (relais de la
sensibilité thermo algique) ; la corne ventrale qui est effectrice, organisée en colonnes qui
s’étendent sur une certaine hauteur de moelle (l’ensemble des colonnes forment la lame IX,
chaque colonne correspond aux corps cellulaire de motoneurones pour un muscle donné, il y a
des colonnes médiales pour les muscles axiaux, des colonnes latérales pour les distaux), ces
colonnes sont entourées par de la substance grise qui forme la lame VII (interneurones), dans
la commissure grise la substance intermédiaire forme la lame X (qui appartient au SN
végétatif), la lame VIII est constitué d’interneurones ; et au niveau thoracique on décrit une
corne latérale.
Les cordons sont des colonnes de substance blanche formées par les axones myélinisés des
voies ascebndantes, descendantes et d’association. On décrit 3 cordons : ventral (entre la
fissure médiane ventrale et le sillon ventral latéral), latéral (entre les sillons ventrolatéral et
dorsolatéral) et dorsal (entre le sillon médian dorsal et le sillon dorso latéral). Les deux
cordons ventraux sont reliés par la commissure blanche ventrale.
Il existe des variations régionales de cette configuration interne. La substance blanche est de
plus en plus importante de bas en haut. Au niveau des segments destinés aux membres, la
substance grise est plus importante ce qui cause 2 renflements. Au niveau du renflement
cervical, les cordons sont très développés surtout le dorsal avec les faisceaux de Goll et
Burdach, et les cornes ventrales et dorsales sont très développées. Au niveau de la moelle
thoracique, il y a peu de substance grise mais il y a des cornes latérales qui sont des centres
végétatifs sympathiques. Au niveau du renflement lombal, il y a peu de substance blanche et
les cornes sont bien développées.
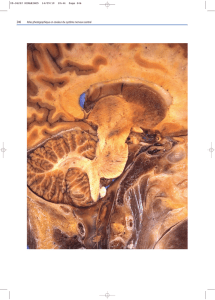
4. Tronc cérébral : anatomie descriptive de la face antérolatérale et émergence des nerfs
crâniens.
Le tronc cérébral est la partie du névraxe situé entre la moelle spinale en bas et le diencéphale
en haut. Il comprend trois parties de bas en haut : la moelle allongée, le pont et le
mésencéphale.
La face antérieure de la moelle allongée présente une fissure médiane ventrale (sillon médian
antérieur) qui prolonge celle de la moelle spinale, de part et d’autre de ce sillon, on trouve les
deux pyramides bulbaires (prolongeant les cordons antérieurs de la moelle) qui sont limitées
en dehors par les sillons latéro-ventraux. Dans la partie supérieure du bulbe, deux saillies

ovoïdes à grand axe vertical, les olives inférieures apparaissent en dehors des pyramides
bulbaires ; à ce niveau le sillon latéral ventral se divise en sillon rétro et pré olivaires.
Au niveau du bulbe : Les nerfs glosso-pharyngiens (IX), vague (X) et le nerf spinal (XI)
émergent (respectivement de haut en bas) entre le sillon latéral ventral et le sillon latéral
dorsal formant le paquet des nerfs mixtes. La douzième paire de nerfs crâniens ou nerf
hypoglosse émerge des sillons préolivaires.
Entre la moelle allongée et le pont se trouve le sillon bulbopontique. Ce sillon devient plus
profond latéralement et forme la fossette latérale, d’où émergent le nerf abducens (VI), le nerf
facial (VII), le nerf intermédiaire (VIIbis) et le nerf vestibulo cochléaire (VIII) formant le
paquet des nerfs stato acoustico facial. Cette région est située au centre de l’angle ponto
cérébelleux.
Le pont qui est le deuxième étage du tronc cérébral présente une volumineuse partie
antérieure traversée par de nombreuses fibres transversales (fibres ponto cérébelleuses). Il est
limité en bas par le sillon médullo pontique et en haut par le sillon ponto pédonculaire qui le
sépare des pédoncules cérébraux. Le sillon basilaire (dans lequel chemine l’artère basilaire)
est une dépression longitudinale sur la ligne médiane de la face antérieure du pont.
Au niveau du tiers supérieur du pont, les racines du nerf trijumeau émergent entre les faces
antérieure et latérale, sa racine sensitive est volumineuse, la racine motrice est petite et
médiale.
Le mésencéphale est limité en bas par le sillon pontopédonculaire, en haut il se continue avec
la jonction mésodiencéphalique, au niveau des tractus optiques.Il est oblique en haut, en avant
et en dehors. Il présente en avant deux volumineux faisceaux blancs ou cru cerebri (pieds des
pédoncules cérébraux). Entre les corps mamillaires (n’appartenant pas au mésencéphale) et
les pédoncules cérébraux latéralement, il existe un espace triangulaire appelé esapce perforé
postérieur répondant à la citerne interpédonculaire. Le nerf oculomoteur (III) naît en dedans
des pédoncules cérébraux au dessus du sillon ponto-pédonculaire.
Sur la partie latérale du tronc cérébral on retrouve les pédoncules cérébelleux rejoignant le
cervelet : on décrit l’inférieur au niveau de la moelle allongée, le moyen au niveau du pont et
le supérieur au niveau du mésencéphale.
Le nerf trochléaire (IV) est le seul à émerger sur la face postérieure du tronc cérébral.
Nerfs crâniens : énumérez : origine, trajet (trous et canaux) et rôle.
5. Nerfs oculomoteurs : III, IV, VI.
Le nerf oculomoteur ou III a pour origine le mésencéphale (son origine apparente en dedans
du pied du crus cerebri ou pied du pédoncule cérébral et son origine réelle en avant de la
substance grise périacqueducale). Il a pour trajet le toit puis la paroi latérale du sinus
caverneux pour passer ensuite dans la fissure orbitaire supérieure, traverse l’anneau de Zinn
où il se traverse en rameaux supérieur et inférieur. Il a pour fonction l’innervation motrice des
muscles droits médial, supérieur, inférieur, oblique inférieur et releveur de la paupière. Il est
viscéro moteur parasympathique pour les muscles intrinsèques (partie annulaire du muscle
ciliaire sphincter de la pupille.), la branche parasympathique du nerf a pour origine réelle le
noyau oculomoteur accessoire et fait relais dans le ganglion ciliaire.
Le nerf trochlaire (IV) a pour origine le mésencéphale (son origine apparente est la face
postérieure du tronc cérébral de part et d’autres de la valvule de Vieussens et son origine
réelle est en avant de la substance grise périacqueducale). Son trajet se fait autour du
mésencéphale, d’abord le toit puis la paroi latérale du sinus caverneux sous le III, pour passer
dans la fissure orbitaire supérieure en dehors de l’annea de Zinn. Son rôle est l’innervation
motrice du muscle oblique supérieur de l’oeil.

Le nerf abducens ou VI a pour origine réelle le pont (relief du colliculus facial et plancher du
V4) et pour origine apparente le sillon bulbo pontique au dessus de la pyramide bulbaire. Il
passe d’abord par le sinus caverneux au dessous de l’artère carotide interne, puis la fissure
orbitaire supérieure dans l’anneau de Zinn. Son rôle est l’inervation motrice du muscle droit
latéral.
6. Nerf trijumeau : V et nerf hypoglosse : XII.
Le nerf trijumeau a pour origine apparente le pont d’où il émerge au tiers supérieur par une
petite racine motrice et une grosse racine sensitive. Ses origines réelles sont un noyau étendu
de la moelle cervicale au mésencéphale (un noyau spinal et sa racine descendante, un noyau
bulbaire, un noyau pontique et un noyau mésencéphlique) et un noyau moteur qui est la
colonne bronchiomotrice pontique. Il se dirige vers le sommet du rocher, pénètre dans le
cavum trigéminal. La racine sensitive donne alors trois branches : ophtalmique (V1),
maxillaire (V2) et mandibulaire (V3), la racine motrice accompagne V3. Son trajet est
différent pour ses trois branches. V1, V2 et V3 naissent du ganglion trigéminal. Le nerf
ophtalmique émerge par la fissure orbitaire supérieure, il est sensitif pour la partie supérieure
de la face (front, dos du nez, cornée, paupière supérieure), pour la muqueuse de la fosse
nasale et des sinus et la duremère des étages supérieur et moyen, il est inclus dans la voie du
réflexe cornéen. Le nerf maxillaire V2 émerge par le foramen rond, il est sensitif pour la joue,
la paupière inférieure, l’aile du nez, la lèvre supérieure, la muqueuse des fosses nasales, des
dents, des gencives supérieures. Le nerf mandibulaire V3 émerge par le foramen ovale, il est
sensitif pour le tiers inférieur de la face (tempe, joue, menton, lèvre inférieure, muqueuse
buccale, les 2/3 antérieures de la langue). La racine motrice accompagne V3 durant son trajet,
elle est motrice pour les muscles masticateurs, tenseur du voile du palais, tenseur du tympan,
mylo-hyoïdien, ventre antérieur du digastrique.
Le nerf hypoglosse a pour origine réelle le plancher du V4 (le noyau de l’hypoglosse se
trouve dans le trigone de l’hypoglosse) et pour origine apparente le sillon préolivaire. Il passe
par le canal de l’hypoglosse, descend dans le cou jusqu’à l’os hyoïde et atteint la langue. Il est
moteur pour l’hémilangue homolatérale.
7. Nerfs mixtes : IX, X, XI.
Le nerf glosso pharyngien, vague et spinal émergent de haut en bas sur la face latérale du
bulbe. Il passe tout les trois par le foramen jugulaire. Le nerf spinal (XI) a une racine spinale
(qui passe par le foramen magnum) et une racine bulbaire qui se rejoignent et passe ensuite
dans le foramen jugulaire. Les nerfs IX et X permettent le carrefour laryngo pharyngé.
Le nerf glosso-pharyngien (IX) est mixte, il est moteur pour le stylo-pharyngien, sensitif pour
le 1/3 postérieur de la langue, le nasopharynx, la trompe d’eustache, la caisse du tympan, le
pharynx buccal, il permet la gustation du tiers postérieur de l’hémilangue homolatérale, il est
végétatif pour la parotide et les barorécepteurs du sinus carotidien. Le nerf vague (X) est
mixte, il est moteur pour la plupart du larynx et du pharynx, il permet la sensibilité du larynx
et du pharynx, la gustation pour la base de la langue et le repli glosso épiglottique, il est
viscéromoteur et sensitif pour le cœur, le poumon, les gros vaisseaux, l’œsophage, l’estomac,
l’intestin grêle, le colon transverse et le sinus carotidien. Le nerf spinal est moteur pour le
sterno cléïdo mastoïdien et le trapèze.

8. Dynamique du liquide céphalorachidien : origine, circulation et résorption
La production du liquide céphalo rachidien se fait au niveau des plexus choroïdes des
ventricules latéraux, de V3 et de V4. Il existe deux compartiments dans la circulation du
LCR : un compartiment intra cérébral formé par les cavités épendymaires (canal central,
aqueduc de Sylvius et les 5 ventricules). Le liquide produit par les plexus choroïdes circule
des VL vers le V3 par les deux foramens interventriculaires (trous de Monro) puis vers le V4
par l’aqueduc de Sylvius et le canal central de la moelle. Dans le toit du V4, le LCR sort de ce
compartiment par l’ouverture médiane (trou de Magendie) pour circuler vers les espaces sous
arachnoïdiens. Les ouvertures latérales sont peu fonctionnelles. Le deuxième compartiment
est le compartiment extra cérébral formé par les espaces sous arachnoïdiens, c’est le lieu de
résorption du LCR au niveau des granulations aracnoïdiennes (de Pacchioni) dans le sinus
longitudinal supérieur et au niveau des trous de conjugaison.
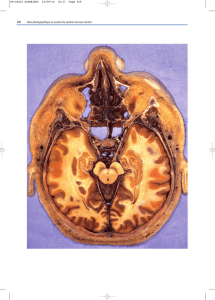
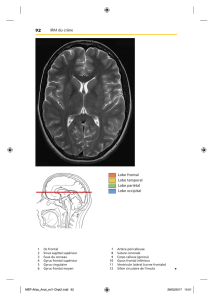
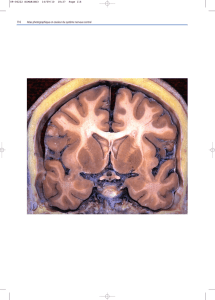
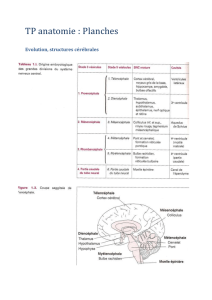
9. Face externe des hémisphères cérébraux : énumérez les principaux sillons et
circonvolutions.
Le cerveau contient 2 hémisphères réunis par des commissures. Il y a trois faces pour chaque
hémisphère : latérale, médiale et inférieure. Son organisation anatomique est basée sur les
sillons. Les sillons profonds délimitent des lobes qui contiennent des circonvolutions ou
gyrus. Sur la face externe ou latérale on peut voir le sillon latéral (scissure de Sylvius) qui
sépare le lobe temporal en bas des lobes pariétal et frontal en haut, le sillon central (scissure
de Rolando) qui sépare le lobe frontal en avant du lobe pariétale en arrière et le sillon pariéto
occipital qui est peu marqué, il sépare le lobe pariétal du lobe occipital.
Le lobe frontal est situé en avant du sillon central, il est subdivisé en gyrus par des sillons
frontaux. Le sillon frontal supérieur se divise en branche ascendante et en branche
descendante, il délimite le gyrus frontal supérieur F1 en haut et le gyrus frontal moyen en bas.
Le sillon frontal inférieur horizontal délimite le gyrus frontal inférieur F3 (lui-même divisé en
3 parties : orbitaire, operculaire ou aire de Broca et triangulaire) en bas. Le sillon précentral
est formé des branches ascendantes et descendantes des sillons frontaux supérieurs et
inférieurs, il limite en avant le gyrus précentral (aire motrice). L’opercule rolandique relie les
gyrus pré et post centraux.
Le lobe pariétal est situé entre le sillon central en avant, le sillon pariéto-occipital en arrière et
le sillon latéral en bas. Il est subdivisé par le sillon intrapariétal se divise en haut en branche
ascendante et en bas en branche descendante (il délimite le gyrus pariétal P1 en haut et le
gyrus pariétal inférieur P2 ) ; et par le sillon post central qui forme la limite postérieure du
gyrus postcentral (qui reçoit toutes les voies de la sensibilité). Le gyrus pariétal inférieur P2
comprend deux circonvolutions situées dans le carrefour pariéto-occipital : le gyrus
supramarginal et le gyrus angulaire qui contourne l’extrémité postérieure du sillon temporal
supérieur.
Le lobe occipital est peu limité en avant par le sillon pariéto occipital et en bas par une
inconstante incisure temporo-occipitale. Il y a donc une continuité temporo occipitale et
pariéto-occipitale. Le sillon occipital supérieur prolonge le sillon intrapariétal, le sillon
occipital inférieur sépare avec le précédent les gyri O1, O2 et O3, le sillon occipital latéral
sépare O2 en partie supérieure et inférieure et atteint en arrière un sillon vertical : le sillon
lunatus.
Le lobe temporal est situé sous le sillon latéral. Il est subdivisé par le sillon temporal supérieur
entre T1 en haut et T2 en bas et par le sillon temporal inférieur entre T2 et T3. Le gyrus
temporal supérieur T1 présente une face supérieure très profonde jusqu’à l’insula : l’opercule
temporal, que l’on peut diviser en régions (planum polare, gyrus temporaux transverses
 6
7
8
9
10
11
12
13
14
15
16
17
6
7
8
9
10
11
12
13
14
15
16
17
1
/
17
100%