Texte

1
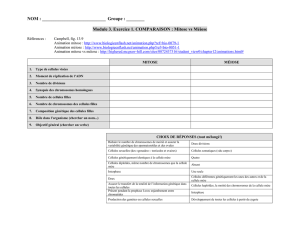
Génétique et gamétogenèse
Rappel sur les chromosomes
Enroulement de l’ADN d’ordre supérieur à la double hélice: nucléosome et chromatine
Chacun des 46 chromosomes humains est composé d’une seule molécule d’ADN double brin.
Etendue, cette molécule ferait environ 2 mètres, mais les cellules possèdent un mécanisme
efficace pour emballer une molécule aussi complexe. Ce système d’empaquetage est d’ailleurs
impliqué dans la régulation de l’expression génétique d’une manière encore mal connue. Des
protéines basiques, les histones, forment une unité de base appelée nucléosome. Le nucléosome
consiste en un noyau central composé de 2 fois 4 molécules d’histones appelées H2A, H2B, H3
et H4. Ces 8 petites protéines très basiques sont ceinturées par 146 paires de nucléotides d’ADN
formant presque deux tours. Les nucléosomes sont séparés par des segments d’ADN, comme les
perles d’un collier. Ce chapelet, d’un diamètre de 10 nm, est encore torsadé pour former la fibre
de chromatine, d’un diamètre de 30 nm. Le niveau le plus élaboré de compaction de l’ADN est le
chromosome.
Chaque espèce possède un assortiment spécifique de chromosomes, appelé caryotype. Le
caryotype humain est de 46 chromosomes. L’homme est un organisme diploïde : 44
chromosomes, appelés autosomes, correspondent à 22 paires de chromosomes homologues. Les
deux autres sont les chromosomes sexuels : les femmes possèdent deux chromosomes X alors
que les hommes possèdent un X et un Y. Les chromosomes humains sont classés de 1 à 22, par
taille décroissante. La plupart d’entre eux sont télocentriques et possèdent un bras court, le bras
« p » (pour “petit”) et un bras long , le bras « q ».

2
La vue classique du chromosome, représenté en mitose, ne correspond pas à son état fonctionnel.
En interphase, lorsque l’expression génique est active, les chromosomes sont déroulés. Ils sont
composés d’une seule fibre de chromatine contenant une double hélice d’ADN et les protéines
associées. Pendant la mitose, les chromosomes subissent un emballage plus dense qui réduit la
longueur des brins de chromatine de 1/10000. Des boucles de chromatine contenant 20-100
kilobase (kb) d’ADN sont attachées à un échaffaudage (scaffold) central composé de protéines
acides, en particulier la topoisomérase II. Ces protéines se lient à des régions riches en
nucléotides AT appelées « scaffold attachment regions ». Dans le chromosome en métaphase,
cet arrangement est encore compacté davantage.
Les chromosomes normaux possèdent un seul centromère, qui apparaît au microscope comme
une zone plus mince et qui est l’endroit où les chromatides sœurs sont appariées. Le centromère
est nécessaire à la ségrégation des chromosomes et les chromosomes qui en sont dépourvus ne
s’attachent pas au fuseau. Lors de l’anaphase, une paire de kinétocores se forment au niveau du
centromère. Des microtubules s’y attachent, raccordant le centromère aux pôles du fuseau.
En anaphase, les microtubules du kinétocore tirent les chromosomes vers les pôles du fuseau.
Les centromères contiennent des séquences d’ADN particulières. Chez les mammifères, ils sont
composés de centaines de kilobases d’ADN répété. Certaines séquences centromériques sont
spécifiques d’un chromosome donné, alors que d’autres sont présentes dans plusieurs
centromères.
Les extrémités des chromosomes sont coiffées de structures complexes composées d’ADN et
de protéines et appelées télomères. La fonction des télomères est de maintenir l’intégrité du
chromosome. En leur absence, le chromosome est instable et fusionne facilement bout à bout
avec des morceaux d’autres chromosome cassés, ou est impliqué dans des phénomènes de
recombinaison, voire dégradé. La présence des télomères est indispensable à la réplication de
l’extrémité des chromosomes. Enfin, les télomères sont probablement fixés à la membrane
nucléaire et servent à maintenir l’architecture des chromosomes dans le noyau en interphase.
Les télomères eucaryotes consistent en longues rangées de répétitions en tandem. Un brin
d’ADN 3’ terminal est riche en TG et apparié au brin 5’ riche en CA. Les séquences
télomériques sont répliquées par un complexe enzymatique spécialisé appelé « télomérase ».

3
En position adjacente des télomères, les chromosomes contiennent également des séquences
particulières, appelées répétitions subtélomériques, dont la fonction est mal connue.
Dans les noyaux en interphase, la plupart de la chromatine est déroulée, dispersée dans le noyau,
et se colore diffusément : elle est appelée euchromatine. Les gènes situés dans l’euchromatine
sont exprimés ou non selon les circonstances. Certains fragments de la chromatine restent
condensés et forment des régions plus densément colorées : c’est l’hétérochromatine. Les gènes
situés dans l’hétérochromatine ne sont pas exprimés. On distingue deux sortes
d’hétérochromatine. L’hétérochromatine constitutive est en permanence condensée et inactive.
Elle contient des séquences hautement répétées et est présente dans et près des centromères
ainsi que dans quelques autres régions. L’hétérochromatine facultative existe sous forme
condensée ou non, comme par exemple dans le cas du chromosome X qui peut être inactivé ou
non. Au sein de l’euchromatine, les bandes G possèdent certaines propriétés de
l’hétérochromatine.
La chromatine y est plus condensée que dans les bandes R et elles sont relativement pauvres en
gènes. La détermination des bandes chromosomiques, en particulier des bandes G, est à la base
d’une nomenclature cytogénétique de localisation chromosomique, les bandes étant nommées
à partir du centromère. L’on dit par exemple que tel locus situé à proximité du centromère du
chromosome 14q est localisé en 14q1.3 (bande1, sous bande 3).
Les anomalies chromosomiques sont variées, relativement fréquentes, et constituent une cause
importante de malformations congénitales et surtout de retard mental. Il n’entre évidemment pas
dans l’objectif de ce cours de les exposer en détails. On distingue les anomalies du nombre des
chromosomes et les anomalies de structure. Parmi les anomalies numériques, les plus fréquentes
sont la trisomie 21 (syndrome de Down, anciennement appelé mongolisme), et les anomalies du
nombre des chromosomes sexuels (X0 : syndrome de Turner ; XXY : syndrome de Klinefelter),
qui seront rappelées dans le chapitre sur la formation des organes génitaux. Parmi les anomalies
de structure, mentionnons le chromosome X fragile, une cause importante de débilité mentale .

4
La mitose et la méiose
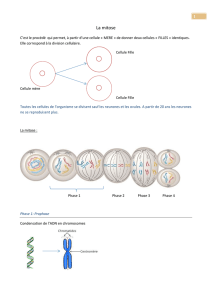
Il existe deux sortes de divisions cellulaires : mitose et méiose. Par la mitose, une cellule
donne naissance à deux cellules génétiquement semblables à elle-même. Par mitose, un seul
oocyte fertilisé donne naissance à un être humain complet composé de 10 exp14 cellules
génétiquement identiques (à quelques exceptions près). Lors de la mitose, chaque cellule fille
reçoit un assortiment complet de 46 chromosomes. La division nucléaire, ou mitose proprement
dite, prend peu de temps. Cependant, elle s’inscrit dans un programme précis, appelé cycle
cellulaire. Après la division, une cellule destinée à se diviser plus avant entre en phase G1, ou
sinon elle entre en phase de repos, appelée G0. Une cellule en phase G1 passe ensuite en phase S
pendant laquelle se produit la réplication semi-conservatrice de l’ADN : chacun des 46
chromosomes est dupliqué en «chromatides sœurs», maintenues assemblées au niveau du
centromère. Après la phase S, la cellule passe en phase G2 puis poursuit en phase M. Lors de la
phase M, les chromatides filles et le centromère deviennent bien visibles et s’alignent selon le
futur plan de clivage de la cellule. Les 46 centromères se divisent et une chromatide de chaque
paire aboutit dans chaque cellule fille.
Méiose et crossing over
La méiose est le type de division cellulaire qui produit les gamètes mâles (spermatozoïde) et
femelles (oocytes), et diffère de la mitose par quelques points très importants. Si le sperme et les
oocytes contenaient un assortiment complet de 46 chromosomes, l’œuf fertilisé en aurait 92. La
méiose sert à réduire le nombre de chromosomes à 23, le nombre haploïde. Elle se compose de
deux divisions appelées méioses I et II. Pendant la méiose I, chaque chromosome se réplique en
deux chromatides sœurs comme dans la mitose. Contrairement à la mitose, cependant, les
chromosomes homologues s’alignent par paires (ce qui est appelé un bivalent
ou synapsis). Chaque bivalent est donc composé de 4 chromatides. Deux chromatides migrent
vers chaque pôle du fuseau en restant attachées. Lors de la méiose II, les chromatides sœurs se

5
séparent, donnant ainsi 23 chromosomes par gamète.
Crossing over
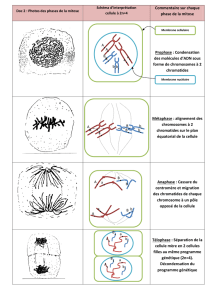
Au cours de la prophase de la méiose I, les brins de chromatine assemblés au sein des bivalents
subissent des échanges plus ou moins aléatoires. Au stade appelé « zygotène », chaque paire de
chromosomes homologues forme un « complexe synaptonemal » où les deux chromosomes sont
accolés et séparés par une zone de protéines. La formation de cette structure marque le début du
stade « pachytène » pendant lequel se produisent les crossing over et la recombinaison. Chaque
crossing over implique une cassure de la double hélice d’ADN dans une chromatide paternelle
et une chromatide maternelle, suivie d’un échange et d’une réparation bout à bout. Le résultat
pratique des crossing over est que chaque chromosome parvenant à un gamète en fin de méiose
contient des segments des deux chromosomes parentaux. Des gènes proches l’un de l’autre sur
le chromosome ont toutes chances de passer ensemble, en bloc, dans les gamètes alors que des
crossing over ont davantage de chances de réassortir des gènes plus éloignés. En moyenne, il se
produit pendant la méiose environ 30-40 crossing over, soit 1 à 2 par chromosome,
approximativement un par
bras chromosomique. Sur le plan morphologique, les crossing over se marquent par la présence
des chiasma. En plus de leur rôle dans la recombinaison, la formation des chiasma semble
indispensable pour permettre la ségrégation des deux chromatides de chaque chromosome
pendant la méiose I. La méiose II se déroule de la même manière que la mitose, sauf qu’il y a 23
chromosomes au lieu de 46. Chaque chromosome est composé de 2 chromatides qui se séparent
en anaphase II. Cependant, alors que les deux chromatides sœurs des chromosomes mitotiques
sont identiques, les chromatides homologues de la méiose II sont différentes en raison des
recombinaisons et crossing-over survenus en méiose I. Lors de la méiose de la femme, les deux
 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%