generalites - LaCaverneD`AliBaba

1
CELLULE
GÉNÉRALITÉS
La cellule constitue l'unité structurelle et fonctionnelle fondamentale des êtres
vivants. Les formes de vie les plus simples sont représentées par des cellules
uniques capables de se reproduire par duplication. Les organismes supérieurs -
c'est-à-dire presque toutes les plantes et les animaux, y compris l'homme - sont
constitués de nombreuses cellules organisées en systèmes complexes qui ont des
fonctions spécialisées et sont reliés entre eux par des systèmes de communication
sophistiqués.
Dès l'Antiquité, un philosophe comme Aristote était parvenu à la conclusion que les
animaux et les plantes, si complexes soient-ils, sont formés de peu d'éléments qui
se répètent dans chacun d'entre eux. Des siècles plus tard, avec l'invention de la
lentille puis du microscope, il a été possible de confirmer ces hypothèses par
l'observation directe.
LA THÉORIE CELLULAIRE
Le mot « cellule » fut utilisé pour la première fois par Robert Hooke (1665), pour
décrire l’unité structurale du liège, à savoir des cavités délimitées par des parois de
cellulose et de suber (liège). Au cours du XIXe siècle, par suite d'un grand nombre
d'observations effectuées aussi bien par des botanistes que par des zoologues, on
vit s'affirmer la théorie cellulaire, selon laquelle tous les organismes - qu’ils soient
simples comme les Bactéries ou complexes comme les plantes et les animaux
supérieurs - sont formés de cellules et de produits cellulaires. Certains d'entre eux
sont formés d'une unique cellule autonome (ce sont les organismes unicellulaires),
tandis que d'autres sont formés de plusieurs cellules différenciées du point de vue
de la forme et des fonctions (ce sont les organismes pluricellulaires).
La théorie cellulaire, qui représente l'une des principales généralisations dans le
domaine biologique, a éclairé toutes les disciplines qui s'occupent de l'étude des
êtres vivants. Une conséquence immédiate de cette théorie a été la constatation
que chaque cellule ne peut que dériver de la division d'une autre cellule. Plus
récemment, grâce aux progrès de la biochimie, on a pu démontrer qu'il existe des
ressemblances fondamentales dans la composition chimique et dans le
métabolisme de toutes les cellules. La théorie cellulaire implique que la fonction de
l'organisme en tant que tout unique est le résultat de la somme des activités et des
interactions des différentes unités cellulaires.
PROCARYOTES ET EUCARYOTES
Le monde des êtres vivants se divise en deux grandes branches selon le type de
cellules qui les constituent. La première branche est celle des Procaryotes (du grec
protos, primitif, et karion, noyau), qui comprend les organismes unicellulaires les
plus simples, c'est-à-dire les Bactéries et les Algues bleues, dépourvus de
membrane nucléaire : le matériel génétique n’est pas enfermé dans un noyau
cellulaire mais est librement immergé dans le cytoplasme. La deuxième branche,

2
celle des Eucaryotes (du grec eu, vrai, et karion, noyau), comprend tous les autres
êtres unicellulaires et pluricellulaires, les végétaux et les animaux, qui ont un noyau
bien défini, avec une membrane qui sépare le matériel génétique du cytoplasme.
Les cellules eucaryotes possèdent en outre une grande variété de compartiments
internes et d'organites chargés d'assurer les fonctions cellulaires.
Tous les organismes, qu’ils soient procaryotes ou eucaryotes, descendent d'une
cellule commune, et se sont différenciés à travers le processus de l'évolution (voir
également évolution des animaux et évolution des plantes).
Les cellules les plus anciennes
Il semble que la vie soit apparue sur la Terre il y a plus de 3 milliards d'années, au
Précambrien inférieur, sous la forme de cellules procaryotes de type bactérien.
Un événement crucial pour la formation de la première cellule a été le
développement d'une membrane externe, c'est-à-dire d'une barrière séparant
quelques macromolécules du milieu environnant. De cette façon, les protéines et
les acides nucléiques, constituants fondamentaux de la cellule vivante, ont pu
interagir dans un espace restreint. La première forme de sélection naturelle se
serait exercée sur ces agrégats de molécules organiques enfermés dans des
compartiments. Les agrégats les plus stables, renfermant des molécules protéiques
ayant une plus grande efficacité enzymatique, auraient été favorisés par rapport
aux agrégats moins stables et peu efficaces.
Les premiers êtres vivants apparus sur la Terre, les Bactéries, représentèrent
l'aboutissement d'une longue phase d'évolution primordiale au cours de laquelle, à
partir de protorganismes renfermés dans une membrane, se seraient développées
et affinées quelques-unes des fonctions premières de la vie, comme la reproduction
et les processus métaboliques.
Les Bactéries sont aujourd'hui encore les organismes les plus simples, présents
dans la plupart des milieux naturels. Elles possèdent une forme sphérique ou en
bâtonnet, et leur longueur est de quelques microns (le micron, ou micromètre, noté
au moyen de la lettre grecque , est une unité de mesure correspondant à un
millième de millimètre). Une épaisse paroi cellulaire renforce la membrane
plasmique des Bactéries, à l'intérieur de laquelle se trouve un compartiment
cytoplasmique sans organisation interne apparente, contenant l'ADN, l'ARN, des
protéines et d'autres molécules. Quand il y a abondance de substances nutritives,
une seule cellule procaryote se divise toutes les 20 minutes, donnant naissance de
la sorte à 5 milliards de cellules (à peu près autant que la population humaine sur la
Terre) en moins de 11 heures. Cette vitesse de reproduction permet aux Bactéries
de s'adapter rapidement aux changements du milieu. En laboratoire, une population
de Bactéries peut évoluer en quelques semaines par mutations spontanées et
sélection naturelle, et devenir ainsi capable d'utiliser de nouveaux types de sucres
(glucides) comme source de carbone.
Il existe des espèces de Bactéries en mesure d'utiliser, pour se nourrir,
pratiquement n'importe quelle molécule organique, y compris les acides aminés, les
graisses (lipides), les polypeptides et les polysaccharides. Une Bactérie placée
dans une solution saline en présence d'une seule source de carbone, telle que le
glucose, doit passer par une série complexe de réactions enzymatiques pour
pouvoir obtenir l'énergie nécessaire aux processus vitaux et à la synthèse des
molécules organiques dont elle a besoin.

3
Au début de la vie sur la Terre, sans doute n'y avait-il pas besoin de tant de
réactions métaboliques. Les Bactéries pouvaient utiliser les molécules organiques
produites antérieurement à l'apparition de la vie sur la Terre, et encore présentes
dans l'océan primordial (soupe primitive). Sous la pression de la sélection naturelle,
et parallèlement à l'épuisement de cette réserve naturelle de substances nutritives,
les Bactéries commencèrent à incorporer le gaz carbonique (CO2) de l'atmosphère
directement dans les composés métaboliques réduits (voir réactions
d'oxydoréduction). En outre, au fur et à mesure que l'ammoniac (NH3) disponible
dans certaines parties du milieu venait à manquer, elles développèrent des voies
métaboliques leur permettant de fixer l'azote atmosphérique (voir cycle de l’azote).
Le développement de la photosynthèse représente une étape fondamentale dans
l'évolution de la vie sur la Terre. C'est en effet grâce à elle que les Bactéries se
transformèrent en organismes autotrophes, c'est-à-dire capables de produire les
composés organiques dont ils ont besoin. Le processus photosynthétique,
toutefois, nécessite que des molécules d'eau (H2O) se scindent pour former des
ions hydrogène (H+). Ce processus a eu pour conséquence la libération de
molécules d'oxygène libre (O2) (voir origine de l'oxygène). L'oxygène atmosphérique
est un composé extrêmement réactif et hautement toxique pour certaines
Bactéries. Avec l'augmentation de l'oxygène dans l'atmosphère, de nombreuses
formes de Bactéries s'éteignirent, tandis que d'autres, telles que les Algues bleues,
firent leur apparition et connurent alors un fort développement.
Les cellules les plus complexes
Le passage des Procaryotes aux Eucaryotes se fit probablement grâce à trois types
différents de Bactéries, qui s’associèrent, commençant à vivre en symbiose (théorie
endosymbiotique).
Les principaux types de Bactéries qui vivaient sur la Terre avant l'apparition de la
cellule eucaryote étaient les Bactéries aérobies, les Bactéries anaérobies et les
Cyanophycées. Les Bactéries aérobies étaient capables d'oxyder l'oxygène (O2) ;
les Bactéries anaérobies pouvaient réaliser la fermentation des substances
organiques avec un rendement énergétique beaucoup plus faible ; les
Cyanobactéries pouvaient quant à elles utiliser la lumière du Soleil pour fabriquer
de façon autonome des substances organiques complexes et riches en énergie à
partir de gaz carbonique (CO2) et d'eau (H2O) (voir métabolisme cellulaire).
L'hypothèse la plus communément admise suppose que la première cellule
eucaryote s'est formée après que de petites Bactéries à respiration aérobie,
apparues depuis peu sur la Terre, pénétrèrent dans une Bactérie anaérobie plus
grande et instaurèrent un rapport de symbiose. Ces deux micro-organismes
auraient tiré des avantages de cette association : les Bactéries aérobies se seraient
trouvées dans un milieu protégé et riche en substances nutritives ; les Bactéries
anaérobies auraient acquis la capacité d'utiliser l'oxygène, devenant
métaboliquement plus actives.
Étant donné l'augmentation continue de l'oxygène dans l'atmosphère, la sélection
naturelle aurait joué en faveur de cette symbiose, qui aurait abouti à la formation
d'une cellule eucaryote primordiale semblable à une amibe. Dans une telle cellule,
les Bactéries aérobies hôtes, transmises lors de la division cellulaire, devinrent les
premières mitochondries. Un deuxième groupe de symbiotes, Bactéries munies de
flagelles, semblables aux spirochètes d'aujourd'hui, se fixèrent à sa surface, lui
donnant la capacité de se déplacer volontairement dans le milieu, et donnant

4
naissance à une sorte d'« amibo-flagellé », ancêtre direct des Champignons et des
animaux. Certaines de ces cellules eucaryotes primitives acquirent un autre
symbiote capable de faire la photosynthèse d’une Cyanobactérie. Ces symbiotes
devinrent des éléments transmissibles par division cellulaire, à la façon des
mitochondries, c’est-à-dire les organites cellulaires appelés les chloroplastes. Ces
cellules à chloroplastes constituent l’origine du règne végétal.
Les cellules eucaryotes, ainsi définies parce que, à la différence ces cellules
procaryotes, elles possèdent un noyau cellulaire, des chloroplastes et des
mitochondries, sont dotées de nombreux autres organites ; elles présentent un
grand nombre de membranes internes. En effet, des membranes nucléaires
entourent le noyau, mais aussi les mitochondries et les chloroplastes. Une série de
membranes forme le réticulum endoplasmique, sorte de compartiment labyrinthique
où sont synthétisés les lipides et les protéines. Un ensemble de poches
membranaires aplaties forment l'appareil de Golgi, système de distribution des
substances élaborées à l'intérieur du réticulum endoplasmique. Les lysosomes
aussi sont limités par des membranes qui empêchent les enzymes qu'ils
contiennent d'attaquer et de détruire les propres protéines de la cellule. De la
même façon, une membrane entoure les péroxysomes, qui contiennent des résidus
métaboliques hautement toxiques pour la cellule.
La quantité de membranes internes d'une cellule eucaryote permet d'augmenter sa
surface d'échange ; elles servent également de substrat pour de nombreuses
réactions métaboliques. Dans une cellule, dont le volume est égal à au moins mille
fois celui de la cellule procaryote, cette augmentation de surface des membranes
est indispensable. Suivant les lois de la géométrie, en effet, le volume augmente en
raison du cube des dimensions linéaires, tandis que la surface augmente
seulement en raison du carré desdites dimensions. Cela veut dire que, si une
grande cellule eucaryote doit maintenir le même rapport surface-volume qu'une
cellule procaryote, il lui faut augmenter sa surface cellulaire au moyen de plis,
d'invaginations et de circonvolutions. Les membranes internes de la cellule
contribuent activement à augmenter la surface de la membrane cellulaire externe
en se fondant avec elle, par les processus d'endocytose et d'exocytose.
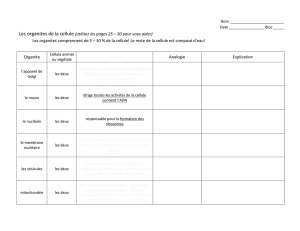
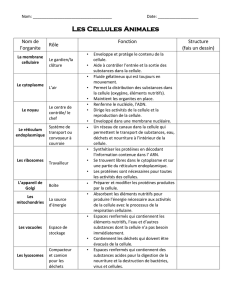
MORPHOLOGIE ET PHYSIOLOGIE DE LA CELLULE
Les cellules sont toutes semblables entre elles : leur diamètre est généralement
compris entre 10 et 30 micromètres, elles sont composées des mêmes molécules
de base, elles les élaborent à travers les mêmes types de réactions chimiques et
elles ont en commun de nombreux organites et des structures identiques ou très
semblables, telles que les membranes, le noyau cellulaire, les ribosomes, l'appareil
de Golgi, le réticulum endoplasmique, les mitochondries et le cytosquelette. Par-
delà cette uniformité de base, il existe des différences entre les cellules animales et
les cellules végétales. Par rapport aux cellules animales, les cellules des plantes
possèdent un revêtement supplémentaire, la paroi cellulaire, formée de cellulose.
Les cellules végétales possèdent en outre des organites caractéristiques : les
vacuoles, vésicules pleines d'eau contenant un certain nombre de substances en
solution, et les chloroplastes, qui contiennent la chlorophylle et d'autres pigments.
L'eau contenue dans les vacuoles confère une certaine rigidité à chaque cellule, et
assure le maintien de la plante. C’est au sein des chloroplastes qu’est capturée
l'énergie du Soleil et qu’elle est transformée en substances énergétiques - les
sucres - grâce au processus de la photosynthèse chlorophyllienne.

5
LA MEMBRANE PLASMIQUE
Chaque cellule est enfermée dans une membrane, une enveloppe protectrice de 8-
12 nanomètres (nm), c'est-à-dire de 8-12 millionièmes de millimètre, qui délimite le
compartiment cellulaire et la sépare du milieu environnant. La membrane joue à la
fois le rôle d'un filtre et d'un moyen de transport. D'une part, elle contrôle l'entrée
des substances nutritives et la sortie des déchets cellulaires et, d'autre part, elle
crée un milieu interne différent du milieu externe. Elle a une autre fonction
importante : celle de créer et de maintenir des concentrations intracellulaires d'ions
spécifiques, c'est-à-dire d'atomes ou de groupes d'atomes portant une charge
électrique. La membrane cellulaire fait également office de capteur des signaux
provenant de l'extérieur, donnant de la sorte à la cellule la possibilité de répondre
aux différents stimuli qu'elle reçoit.
Les constituants principaux de la membrane plasmique sont les lipides (c'est-à-dire
les graisses), les protéines et, dans certains cas, les hydrates de carbone (ou
sucres, appelés aussi glucides).
Les lipides sont les constituants fondamentaux de la membrane ; ils sont
représentés principalement par les phospholipides, les glycolipides et les stérols.
Les phospholipides sont des lipides complexes caractérisés par une tête polaire
hydrophile (c'est-à-dire une extrémité portant une charge électrique négative
soluble dans l'eau), et de longues queues apolaires hydrophobes (lesquelles,
n'ayant pas de charge électrique, ne se mélangent pas avec l'eau). Ces
caractéristiques moléculaires font que les phospholipides, plongés dans une
solution aqueuse, forment une bicouche fluide dans laquelle les têtes hydrophiles
entrent en contact avec l'eau, tandis que les longues queues hydrophobes se
disposent vers l'intérieur, s'isolant du milieu aqueux. Cette bicouche lipidique de
5 nanomètres d'épaisseur sert de barrière presque imperméable au passage de
substances solubles dans l'eau. Les glycolipides, tout comme les phospholipides,
sont des lipides complexes dotés de têtes hydrophiles et de queues hydrophobes,
tandis que les stérols sont complètement apolaires.
Les hydrates de carbone sont présents dans la membrane sous la forme
d'oligosaccharides, courtes chaînes formées par l'association de quelques
molécules de sucres simples. Ces chaînes sont à leur tour liées à des protéines ou
des lipides de membrane, formant respectivement des glycoprotéines et des
glycolipides.
Dans la bicouche lipidique sont immergées des molécules de protéines qui, grâce à
la fluidité qui caractérise la membrane, peuvent changer de position en se
déplaçant latéralement. Ces protéines (protéines intrinsèques) pénètrent
partiellement ou bien traversent toute l’épaisseur de la membrane, et débordent sur
l’une ou sur les deux surfaces, externe et interne. D’autres protéines, les protéines
extrinsèques, sont dispersées sur la surface externe ou sur la surface interne de la
membrane. Les protéines de membrane ont une fonction bien déterminée.
Certaines transportent des substances spécifiques à l’intérieur et à l’extérieur de la
cellule, d’autres forment des canaux ou des pores à travers lesquels peuvent
passer les molécules polaires pour lesquelles la double couche lipidique constitue
une barrière. Ils existe d’autres types de protéines tout aussi importantes : celles
qui fonctionnent comme des récepteurs, autrement dit comme des sites spécifiques
auxquels se lient des substances particulières comme les hormones, et celles qui
jouent le rôle de catalyseurs de certaines réactions enzymatiques.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
1
/
46
100%