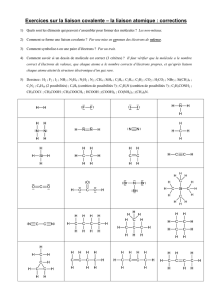
TP word exercice 2009 brut

Métabolisme II:
Flux d'énergie dans la cellule
I.Respiration cellulaire aérobie
I.1.Vue d'ensemble
I.1.a.Équation générale
Le principe de la respiration cellulaire aérobie est relativement similaire à celui d'un moteur de
voiture. Du glucose réagit avec de l'oxygène pour former du dioxyde de carbone et de l'eau et
libérer de l'énergie.
La dégradation du glucose est exergonique (∆G = -2871 kJ/mol). L'énergie libérée sous forme
d'ATP va participer au travail cellulaire.
I.1.b.Réactions d'oxydo-réduction
Pourquoi les voies cataboliques fournissent-elles de l'énergie lors de la dégradation du glucose ? Le
transfert des électrons qui se produit durant les réactions chimiques libère de l'énergie emmagasinée
dans les molécules de nutriments. Cette énergie sert à synthétiser de l'ATP.
Le transfert d'électrons d'un réactif à un autre au cours d'une réaction chimique est appelé réaction
d'oxydo-réduction. L'oxydation correspond à la perte d'électrons; la réduction à un gain d'électrons.
Un agent réducteur est une substance qui donne des électrons tandis qu'un agent oxydant est une
substance qui en accepte.
Une réaction d'oxydo-réduction qui rapproche des électrons de l'oxygène, O2 (fort électronégatif),
libère de l'énergie chimique.
La combustion de l'essence dans un moteur est une réaction d'oxydo-réduction; l'énergie libérée
actionne les pistons et le véhicule avance. La respiration cellulaire est une réaction d'oxydo-
réduction au cours de laquelle le glucose est oxydé et l'oxygène est réduit.
Les liaisons des molécules organiques riches en hydrogène renferment des électrons à énergie
potentielle élevée susceptibles de se rapprocher de l'oxygène. Lors de la respiration cellulaire,
l'hydrogène du glucose est transféré à l'oxygène. En effet, la perte d'électrons est souvent associée à
la perte d'un proton.
I.1.c.Libération progressive de l'énergie
Si l'énergie de la combustion du glucose était libérée en une fois, il serait difficile de l'exploiter. En
effet, la libération en un bloc de l'énergie de l'essence (explosion) ne fait pas fonctionner le moteur.
La respiration cellulaire se produit en une série d'étapes catalysées par des enzymes. Lors de
certaines étapes, des atomes d'hydrogène sont arrachés au glucose mais ne vont pas directement se
lier à l'oxygène; ils sont transférés sur une coenzyme, le nicotinamide adénine nucléotide (NAD+).
Des enzymes, les déshydrogénases retirent une paire d'atomes d'hydrogène du substrat, soit deux
électrons et deux protons (H+). Elles apportent ensuite les deux électrons et un proton au NAD+; le
second proton est libéré dans la solution environnante.
Les électrons perdent très peu d'énergie lors de leur transfert sur le NAD+. Par conséquent, chaque
mole de NADH+H+ produit durant la respiration représente une réserve d'énergie pour la
production de l'ATP. Le NADH va apporter les électrons (énergétiques) au sommet de la chaine de
transport des électrons; au bas de cette chaine se trouve l'oxygène, accepteur final des électrons.
Le transfert des électrons du NADH+H+ à l'oxygène est une réaction exergonique (∆G = -222
kJ/mol). Toutefois, cette énergie n'est pas libérée en une fois mais progressivement car les électrons
descendent la chaine en passant d'un transporteur à un autre jusqu'à atteindre l'oxygène, an perdant
un peu d'énergie à chaque étape.
I.1.d.La mitochondrie, siège de la respiration cellulaire aérobie
La mitochondrie est un organite cellulaire présent dans presque toutes les cellules eucaryotes; elle
est le siège de la respiration cellulaire. Elle mesure entre 1 et 10 µm de long et leur nombre dans la
cellule dépend de l'activité métabolique.
La mitochondrie est entourée d'une enveloppe faite de deux membranes: la membrane
mitochondriale externe et la membrane mitochondriale interne. La membrane mitochondriale
interne est repliée sur elle-même de façon à former de nombreuses crêtes et divise la mitochondrie
en deux compartiments: l'espace intermembranaire et la matrice mitochondriale.

La mitochondrie possède son propre ADN.
I.1.e.Déroulement
La respiration cellulaire est divisée en trois stades principaux: la glycolyse, le cycle de Krebs et la
chaine de transport des électrons.
Durant les deux premiers stades, il y a dégradation du glucose (d'abord en pyruvate durant la
glycolyse puis en gaz carbonique durant le cycle de Krebs). La chaine de transport des électrons,
troisième stade, accepte les électrons provenant des produits des deux premières étapes par
l'intermédiaire du NADH+H+ et du FADH2. A la fin de la chaine, les électrons se combinent à des
protons et de l'oxygène (O2) pour former de l'eau. Durant la descente de cette chaine, les électrons
libèrent de l'énergie qui est utilisée pour former de l'ATP par phosphorylation oxydative.
De l'ATP est également formé durant certaines étapes de la glycolyse et du cycle de Krebs par un
mécanisme appelé phosphorylation au niveau du substrat. Il y a, dans ce cas, transfert d'un
phosphate d'un substrat à de l'ADP pour former de l'ATP.
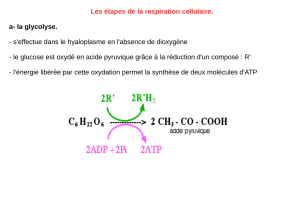
I.2.Glycolyse
I.2.a.Généralités
La glycolyse est la première phase de la respiration cellulaire. Cette voie catabolique de dégradation
du glucose se retrouve dans presque toutes les cellules vivantes. La glycolyse se déroule dans le
cytosol de la cellule et ne nécessite pas l'apport d'oxygène.
Durant cette phase, le glucose (monosaccharide à six atomes de carbone) est scindé en deux
pyruvates (monosaccharide ayant trois atomes de carbone).
I.2.b.Phase 1: phase d'investissement en énergie
Durant cette première phase, il y a phosphorylation du glucose en fructose-1,6-biphosphate. Cette
molécule instable se scinde alors en deux molécules de glycéraldéhyde-3-phosphate (ou
phosphoglycéraldéhyde).
Pour former ces deux molécules à partir de glucose, il y a consommation de deux molécules d'ATP.
I.2.c.Phase 2: phase de libération d'énergie
Au cours de cette seconde phase de la glycolyse, chaque molécule de phosphoglycéraldéhyde est
convertie en une molécule de pyruvate. Cette conversion libère 1 H2O, 1 NADH+H+ et 2 ATP par
pyruvate produit, soit 2 H2O, 2 NADH + 2 H+ et 4 ATP par glucose dégradé.
I.2.d.Bilan
Au cours de la glycolyse, par mole glucose dégradée, sont produits deux moles de pyruvate, deux
moles d'H2O, deux moles d'ATP et deux NADH + 2 H+. Les molécules d'ATP formée durant la
glycolyse le sont par phosphorylation au niveau du substrat.
I.3.Cycle de Krebs
Le cycle de Krebs, ou cycle des acides tricarboxyliques ou encore cycle de l'acide citrique, est la
deuxième étape de la respiration cellulaire; il a lieu dans la matrice mitochondriale. Toutefois, il
intervient également dans de nombreuses autres voies métaboliques (catabolisme des acides gras,
des acides aminés,... fabrication du glucose, du cholestérol, ...). La glycolyse a libéré moins du quart
de l'énergie contenue dans le glucose; le reste est stocké dans les deux moles de pyruvate.
I.3.a.Conversion du pyruvate
Le pyruvate, produit lors de la glycolyse, entre dans la mitochondrie (matrice mitochondriale) où il
est converti en acétyl-CoA. La transformation du pyruvate en acétyl-CoA est une réaction d'oxydo-
réduction qui produit une mole de CO2 et une mole de NADH+H+ par mole de pyruvate
transformée.
L'acétyl-CoA est un complexe composé d'acétate (issu du pyruvate ayant perdu son groupement
carboxyl) et de la coenzyme A (dérivé de la vitamine B).
I.3.b.Cycle proprement dit
L'acétyl-CoA entre dans le cycle de Krebs en se liant à de l'oxaloacétate pour former du citrate. Ce
dernier est alors progressivement dégradé en oxaloacétate, libérant deux moles de CO2, 3 NADH +
3 H+, une mole d'ATP ainsi qu'un FADH2 (la flavine adénine dinucléotide, composé dérivé de la
vitamine B) par mole de pyruvate.
I.3.c.Bilan

Au cours de la seconde étape de de la respiration cellulaire sont libérés trois moles de CO2, quatre
moles de NADH, quatre protons, une mole de FADH2 et une mole d'ATP par mole de pyruvate.
I.4.Chaine de transport des électrons et phosphorylation oxydative
Les molécules de NADH et de FADH2, engendrées par la glycolyse et le cycle de Krebs,
transmettent leurs électrons à la chaine de transport des électrons. Ce sont eux qui vont libérer la
majeure partie de l'énergie du glucose.
I.4.a.Chaine de transport des électrons
La chaine de transport des électrons est constituées d'un ensemble de molécules (la plupart des
protéines) enchassées dans la membrane mitochondriale interne. Grâce aux crêtes qui augmentent
sa surface, cette membrane contient des milliers d'exemplaires de la chaine.
Le NADH + H+ cède ses électrons au premier transporteur de la chaine, une flavoproteine (FMN).
L'arrivée d'électrons réduit cette protéine qui, pour retrouver sa forme oxydée, cède à son tour les
électrons au second transporteur. Les électrons passent ainsi d'un transporteur à l'autre en perdant
progressivement de l'énergie libre. Le dernier transporteur de la chaine donne ses électrons à
l'oxygène (O2) qui recueille également une paire de protons du milieu pour former de l'eau.
Le FADH2 cède également ses électrons au niveau de la chaine de transport mais directement au
troisième transporteur.
La chaine de transport des électrons ne produit pas directement de l'ATP.
I.4.b.Chimiosmose et formation d'ATP
L'énergie libérée par les électrons lors de leur descente de la chaine de transport est utilisée pour
véhiculer les protons (H+) à travers la membrane mitochondriale interne, depuis la matrice vers
l'espace intermembranaire. Les protons ont alors tendance à refluer vers la matrice mitochondriale,
à cause d'un gradient électrochimique.
Le retour des protons vers la matrice mitochondriale ne peut se faire qu'à travers un canal situé au
cœur d'une ATP synthétase. Ce complexe protéique entraine une phosphorylation oxydative de
l'ADP en ATP, alimentée par le retour exergonique des protons. Ce mécanisme est appelé
chimiosmose (poussée des protons à travers la membrane).
L'ATP synthétase est constituée de trois parties: le rotor (inséré dans la membrane mitochondriale
interne qui tourne à cause du passage des électrons, l'arbre (suivant les mouvements du rotor, il
active les sites catalytiques du stator) et le stator (immobile et situé dans la matrice, il contient les
sites catalytiques qui combinent l'ADP et le phosphate inorganique pour former l'ATP).
I.4.c.Bilan
Les électrons transférés à la chaine de transport des électrons génèrent (indirectement) trois moles
d'ATP s'ils sont issus d'une mole de NADH et deux moles d'ATP s'ils proviennent d'une mole de
FADH2.
I.5.Bilan énergétique de la respiration cellulaire aérobie
Au total, au cours de la respiration cellulaire, 38 moles d'ATP sont produites par mole de glucose
dégradée.
II.Fermentation
II.1.Principe
En absence d'oxygène, certaines cellules utilisent la fermentation pour oxyder leurs nutriments et
fabriquer de l'ATP.
Lors de la glycolyse (étape ne nécessitant pas d'oxygène) une mole glucose est oxydée en deux
moles de pyruvate. L'agent oxydant, c'est-à-dire la substance qui accepte les électrons est le NAD+.
Durant cette étape, il y a production de deux moles d'ATP par mole de glucose par phosphorylation
au niveau du substrat.
La fermentation est un prolongement de la glycolyse. En effet, pour fabriquer de l'ATP, il faut qu'il
y ait suffisamment de NAD+ dans la cellule. Donc, sans un mécanisme de recyclage du NADH +
H+, la cellule épuiserait rapidement toutes ses réserves de NAD+. Dans le cas de la fermentation,
les électrons du NADH + H+ sont transférés au pyruvate (produit de la glycolyse) ou à un de ses
dérivés; du NAD+ est ainsi de nouveau prêt à accepter des électrons issus de l'oxydation du glucose.
Il existe plusieurs types de fermentation dont la fermentation alcoolique et la fermentation lactique.

II.2.Fermentation lactique
Au cours de la fermentation lactique, le pyruvate se fait réduire directement par le NADH+ H+. Du
lactate est ainsi formé.
Ce mode de fermentation (chez des bactéries et des levures) est utilisé dans l'industrie laitière pour
la fabrication de fromages et yogourt.
Chez l'homme, l'accumulation du lactate dans les muscles au cours d'un exercice exigeant (le
glucose se fait dégrader plus vite que l'arrivée d'oxygène; donc la fermentation commence)
provoque de la fatigue et des douleurs. Ensuite, le lactate est transporté au niveau du foie où il est
transformé en pyruvate.
II.3.Fermentation alcoolique
Au cours de la fermentation alcoolique, le pyruvate (issu de la glycolyse) est converti en éthanol en
deux étapes. Le pyruvate est d'abord transformé en acétaldéhyde par retrait de dioxyde de carbone.
L'acétaldéhyde est alors réduit en éthanol par l'apport d'électrons provenant du NADH+ H+.
Dans l'industrie brassicole ou vinicole, la fermentation alcoolique est déclenchée au moyen de
levures. Beaucoup de bactéries réalisent aussi la fermentation alcoolique.
II.4.Bilan de la fermentation
Durant la fermentation, alcoolique ou lactique, la seule étape productrice d'énergie est la glycolyse.
Au cours de cette étape, la dégradation d'une mole de glucose permet la fabrication de deux moles
d'ATP.
III.Photosynthèse
La photosynthèse a lieu chez les végétaux chlorophylliens. Elle s'observe également chez les algues
et chez certaines bactéries (dont les cyanobactéries et les bactéries pourpres sulfureuses). Elle
permet de produire des molécules organiques (glucides) nécessaires aux végétaux à partir de
dioxyde de carbone (CO2) et d'eau (H2O). Cette production de molécules organiques nécessite de
l'énergie sous forme de lumière.
III.1.La photosynthèse se déroule dans les chloroplastes
Chez les végétaux supérieurs, la photosynthèse se déroule dans les chloroplastes (organites
cellulaires d'environ 5 µm de long) au niveau des thylakoïdes. Il s'agit de sacs aplatis limités par une
membrane et contenant des molécules de pigments. L'espace intérieur des thylakoïdes est appelé
espace intrathylakoïdien.
Dans certaines régions du chloroplaste, les thylakoïdes s'empilent pour former des structures
appelées grana. Les thylakoïdes baignent dans un liquide appelé le stroma.
Toutes les cellules vertes d'une plante contient des chloroplastes mais se sont les cellules des
feuilles qui en contiennent le plus; un demi millions par mm² de feuille.
Comme les mitochondries, les chloroplastes ont leur propre ADN, leurs ARN et leurs ribosomes.
III.2.Aspects généraux de la photosynthèse
Durant la photosynthèse, du gaz carbonique et de l'eau sont consommés pour produire du glucose et
de l'oxygène. Cette réaction est endergonique, elle demande de l'énergie. Celle-ci est fournie par la
lumière.
La photosynthèse se déroule en deux phases. La première phase correspond à la transformation de
l'énergie lumineuse en énergie chimique grâce à des réactions appelées réactions photochimiques.
Cette première phase est parfois appelée phase claire ou phase lumineuse car elle nécessite l'apport
de lumière. L'absorption de lumière provoque un transfert d'électrons et de protons de l'eau vers un
accepteur, le NADP+ (nicotinamide adénine dinucléotide phosphate) qui devient alors du NADPH
+ H+. Il y a également production d'ATP par photophosphorylation.
L'énergie chimique produite (ATP) va ensuite être utilisée lors de la seconde phase (cycle de
Calvin). Ce cycle correspond à une série de réactions chimiques qui vont permettre la
transformation progressive du gaz carbonique (CO2) en glucides (qui pourront être utilisés par le
végétal). La seconde phase est parfois nommée phase sombre ou phase obscure car elle ne nécessite
pas de lumière. Au cours de cette étape, du CO2 est incorporé puis réduit en glucide par l'ajout
d'électrons en provenance du NADPH + H+.
III.3.Réactions photochimiques

Les réactions photochimiques, première phase de la photosynthèse, se déroulent dans les
thylakoïdes des chloroplastes.
III.3.a.Pigments
Les pigments sont des substances qui absorbent la lumière visible chez les organismes
photoautotrophes. Chaque pigment absorbe surtout certaines longueurs d'onde de la lumière et les
fait ainsi disparaitre. La partie de la lumière qui n'est pas absorbée est diffusée; c'est dans cette
couleur qu'apparait à l'œil humain l'objet contenant le pigment. La capacité d'absorption d'un
pigment en fonction de la longueur d'onde de la lumière est le spectre d'absorption du pigment.
Les chloroplastes possèdent deux pigments principaux, la chlorophylle a et la chlorophylle b, ainsi
que quelques pigments accessoires, les pigments caroténoïdes. Le spectre d'absorption de la
chlorophylle a révèle que la lumière bleue/mauve et la lumière rouge sont les plus favorables à la
photosynthèse. La chlorophylle b absorbe la lumière bleue et la lumière orange. Les caroténoïdes
absorbent dans les lumières bleues-mauves.
Aucun des pigments de la photosynthèse n'absorbe la lumière verte; elle est donc diffusée. C'est
pourquoi, les feuilles (et autres parties chlorophylliennes) des végétaux apparaissent vertes.
III.3.b.Photo-oxydation de la chlorophylle
La lumière pénètre dans les cellules jusqu'aux chloroplastes et heurte les amas de pigments présents
au niveau de la membrane des thylakoïdes.
Quand une molécule de chlorophylle absorbe un photon, un de ses électrons passe à une orbitale où
il possède davantage d'énergie potentielle. Le pigment passe ainsi de l'état fondamental à l'état
excité. L'état excité est instable; l'électron revient donc à l'état fondamental (en 10-9s) et libère le
surplus d'énergie sous forme de chaleur et d'un photon (qui va être capté par un pigment ...).
L'émission de photons par les pigments est appelée fluorescence.
Les pigments sont insérés dans la membrane des thylakoïdes de chaque chloroplaste sous forme de
complexes appelés photosystèmes. Chaque photosystème comporte entre 250 et 400 molécules de
pigment ainsi que des protéines et autres petites molécules organiques. Au sein d'un photosystème,
la plupart des pigments sont capables d'absorber l'énergie lumineuse; ce sont les pigments
antennaires. Toutefois, seule une molécule de chlorophylle a, située au cœur du photosystème, peut
utiliser cette énergie pour déclencher la photosynthèse. Cette molécule de chlorophylle a est
associée à un accepteur primaire d'électrons pour former le centre réactionnel.
Quand la lumière frappe un pigment antennaire, l'énergie (sous forme de photons) est transmise de
proche en proche jusqu'au centre réactionnel. La chlorophylle a du centre réactionnel est excitée.
L'électron excité de la chlorophylle a est alors recueilli par l'accepteur primaire avant son retour à
l'état fondamental. C'est la première étape des réactions photochimiques.
Il existe deux types de photosystèmes : les photosystèmes I et les photosystèmes II (numérotés dans
l'ordre de leur découverte). Ils diffèrent par leur centre réactionnel.
III.3.c.Photosystème I
La lumière absorbée par les pigments antennaires provoque l'excitation d'électrons puis la libération
d'énergie sous forme de photons. Cette énergie est captée par le pigment voisin où le même
phénomène se produit. Progressivement l'énergie atteint le centre réactionnel du photosystème I.
Deux électrons excités du photosystème I sont captés par l'accepteur primaire. La chlorophylle a
(centre réactionnel) a donc perdu deux électrons.
Les deux électrons énergétiques, captés par l'accepteur primaire, passent au travers d'une chaîne de
transport des électrons. Le dernier accepteur d'électrons de cette chaîne est le NADP+. En acceptant
les deux électrons et en s'associant à deux protons, le NADP+ est converti en NADPH + H+. Ce-
dernier est un agent réducteur nécessaire à la production des glucides durant le cycle de Calvin.
Pour rétablir l'équilibre électronique, il est nécessaire de remplacer les deux électrons manquant au
niveau de la chlorophylle a du centre réactionnel du photosystème I. Pour ce faire, deux électrons
viennent depuis le photosystème II.
III.3.d.Photosystème II
De la même manière que pour le photosystème I, le photosystème II absorbe de l'énergie lumineuse
au niveau des pigments antennaires. Cette énergie est progressivement transmise au centre
 6
6
1
/
6
100%