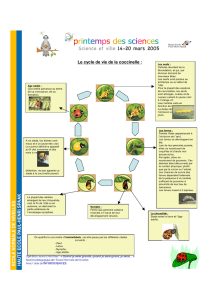
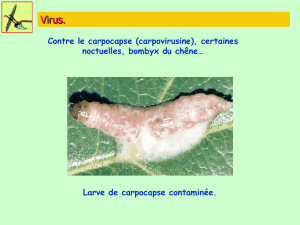
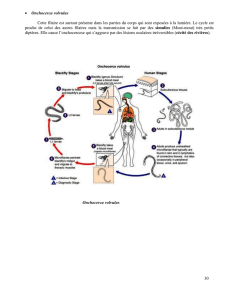
Extrait de la These_J-P. SARTHOU, 1996

Synthèse bibliographique sur les Syrphidés – Thèse de Jean-Pierre SARTHOU 1996
1
AVERTISSEMENT : cette synthèse bibliographique date de plusieurs années et n’est plus à
jour sur quelques points notamment systématiques. Exemple : il y est dit que « la clé de
Séguy 1961 est encore valable pour le genre Chrysotoxum, à quelques changements de
nomenclature près », ce qui n’est pas le cas aujourd’hui et aucune clé publiée n’est
totalement satisfaisante pour ce genre. On consultera avec profit la base de données Syrph
the Net annuellement mise à jour pour tous ces aspects systématiques.
Néanmoins, la grande majorité des informations bioécologiques sont évidemment toujours
valables même si de nouvelles connaissances ont été apportées depuis.
CHAPITRE II - LES DIPTERES SYRPHIDAE COMME MODELE D’ETUDE
I - Introduction :
Pour tenter de mener à bien une telle étude agroécologique portant sur les aspects de préservation
de la biodiversité dans les agroécosystèmes et d’amélioration de l’activité des organismes auxiliaires
des cultures, il est nécessaire de choisir judicieusement un taxon adéquat.
Ce dernier doit, en premier lieu, renfermer suffisamment d’espèces dans la zone biogéographique
considérée, espèces dites “indicatrices de l’environnement” en occupant la plus large gamme
possible de biotopes afin de bien orienter les études de biologie de la conservation et les choix de
gestion de milieux. Il doit, en deuxième lieu, présenter une systématique relativement bien établie,
donnant un minimum de problèmes d’identification aux spécialistes. En effet, "seules des bases
faunistiques et systématiques solides peuvent permettre de progresser dans une étude écologique
sérieuse" (GOELDLIN DE TIEFENAU, 1974). Enfin, le taxon doit posséder un certain nombre d’espèces
jouant un rôle bénéfique significatif au sein des cultures.
Le taxon retenu dans cette étude pour toutes ces raisons, occupe le rang de la famille : il s’agit de
celle des Syrphidae (Diptères). Ce sont d’élégants Diptères, recherchant les fleurs au stade imaginal,
prédateurs, phytophages ou saprophages au stade larvaire, bien connus des jardiniers, promeneurs
et agriculteurs un tant soit peu observateurs car facilement reconnaissables à leur vol caractéristique,
rapide, parfois riche en circonvolutions aériennes et entrecoupé de phases stationnaires, et à leur
livrée souvent sombre à taches ou bandes claires leur conférant un habitus mimétique de certains
Hyménoptères.
Les Syrphidae sont de bons insectes marqueurs de l’environnement car satisfaisant à la plupart des
critères énoncés par SPEIGHT (1986a) : moins de 5% des espèces posent des problèmes sérieux
d’identification aux spécialistes, la plupart des espèces ont des exigences écologiques connues dans
leurs grandes lignes ou avec précision et plusieurs espèces peuvent permettre de mettre en
évidence des caractéristiques de biotopes indécelables avec des vertébrés ou des plantes
supérieures ; enfin, de plus en plus de listes d’espèces concernant diverses contrées d’Europe sont
publiées. Ces Diptères sont de plus de précieux auxiliaires de l’agriculture grâce aux adultes
floricoles participant activement à la pollinisation des cultures et surtout grâce aux larves de
nombreuses espèces (sous-famille des Syrphinae et tribu des Pipizini) qui ont un régime zoophage
et particulièrement aphidiphage, donc grandes consommatrices de pucerons y compris ceux des
cultures pour certaines d’entre elles.

Synthèse bibliographique sur les Syrphidés – Thèse de Jean-Pierre SARTHOU 1996
2
II - Position systématique :
La famille des Syrphidae fait partie de l’ordre des Diptères, Invertébrés de l’embranchement des
Arthropodes, du sous-embranchement des Mandibulates, de la classe des Insectes, de la sous-
classe des Ptérygotes, du groupe des Holométaboles et du super-ordre des Mécoptéroïdes.
Parmi les Diptères, les Syrphidae se classent dans le sous-ordre des Cyclorrhaphes, dans le groupe
des Aschiziens et dans la super-famille des Syrphidoidea avec les Pipunculidae.
L'ouvrage "Diptères Syrphides de l’Europe occidentale" de E. SEGUY (1961), bien que toujours utile à
consulter ponctuellement (pour le genre Chrysotoxum par exemple, après avoir tenu compte des
changements récents de nomenclature), est selon les propres termes de l’auteur "loin d’être
exhaustif" et a été de plus réalisé à une période où plusieurs genres nécessitaient une révision au
niveau européen. Ainsi, après plus de trente ans, cet ouvrage de systématique et d’identification (le
dernier d’édition française) ne suffit plus et il est nécessaire de se tourner aujourd’hui vers des
travaux étrangers rassemblés dans une bibliographie malheureusement complètement disséminée,
ce qui est un sérieux handicap au commencement de la recherche.
La systématique infra-famille adoptée dans le présent travail suit celle de VOCKEROTH (1969) et de
PECK (1988) concernant les trois sous-familles : Syrphinae, Milesiinae et Microdontinae. Nous
incluons ici le genre Microdon dans la sous-famille des Microdontinae et donc dans la famille des
Syrphidae, simplement par souci de commodité et pour ne pas omettre ses espèces toujours
incluses dans cette sous-famille par de nombreux auteurs. SPEIGHT (1993, 1994) suggère, d'après les
études morphologiques externes des adultes (SPEIGHT, 1987), des larves (HEISS, 1938 ; TORP, 1984)
et suite au fait que les adultes n'aient jamais été vus se nourrissant de pollen ou de nectar (DE BUCK,
1990), qu'il en soit fait une famille à part entière. Nous approuvons les suggestions de SPEIGHT et
rappelons que nous ne procédons à ce regroupement que par commodité et resignalerons quand
nécessaire l'appartenance de ce genre à la famille des Microdontidae.
Concernant la subdivision en tribus et genres, nous suivons les propositions de VOCKEROTH (1969),
THOMPSON et al. (1982), VOCKEROTH (1986), VOCKEROTH & THOMPSON (1987), GOELDLIN DE TIEFENAU
(1989), ROTHERAY & GILBERT (1989), GOELDLIN DE TIEFENAU et al. (1990) et VOCKEROTH (1990, 1992)
pour les Syrphinae.
Concernant les Milesiinae, nous avons essentiellement suivi HIPPA (1978a, 1978b), THOMPSON et al.
(1982), MAIBACH & GOELDLIN DE TIEFENAU (1994) et MAIBACH et al. (1994a, 1994b).
Les classifications et nomenclatures génériques et spécifiques des deux sous-familles diffèrent
néanmoins sur quelques points, car elles suivent GOELDLIN DE TIEFENAU (1989), MAIBACH et al.
(1992), SPEIGHT & LUCAS (1992), SPEIGHT (1993, 1994), SPEIGHT et al. (en préparation) et SARTHOU &
GOELDLIN DE TIEFENAU (en préparation) :
les trois entités suivantes acquièrent un statut générique propre parmi les Syrphinae :
Epistrophella, Meligramma, Olbiosyrphus
de même pour Anasimyia, Helophilus et Parhelophilus pour les Milesiinae
Megasyrphus annulipes auct. nec.(Zett.) devient Eriozona erratica (L.)
Chrysotoxum festivum auct. nec. L. devient C. arcuatum L.
Chrysotoxum arcuatum auct. nec. L. devient C. fasciatum (Müller)
Sphaerophoria menthastri auct. nec. L. devient S. interrupta (Fabricius)
Sphaerophoria menthastri L. nouveau synonyme de S. scripta (L.)
Doros conopseus auct. nec. Fabricius devient D. profuges (Harris)
Pipizella varipes auct. nec. (Meigen) devient P. viduata (L.)
Paragus bradescui revalidé au rang de bonne espèce.
Contrairement à d'autres familles de Diptères, les Syrphidae, qui comptent parmi les groupes les plus
difficiles de l'ordre, ont fait l'objet d'importants travaux de recherche en systématique ces quatre
dernières décennies notamment. Toutefois, des problèmes taxinomiques significatifs demeurent.
Dans la sous-famille des Milesiinae, trois genres posent toujours des problèmes quant à
l'identification de leurs espèces : Cheilosia, Eumerus et Merodon (SPEIGHT, 1993) et de sérieux

Synthèse bibliographique sur les Syrphidés – Thèse de Jean-Pierre SARTHOU 1996
3
problèmes d'identification d'espèces persistent pour d'autres genres ; le genre Pipiza notamment ne
permet pas aujourd'hui encore de délimiter la variabilité de chacune de ses espèces avec
suffisamment de confiance (SPEIGHT, 1993) ; il est également impossible actuellement d'identifier les
femelles du genre Pipizella Rondani et des espèces brevidens (Egger), latitarsis (Egger), pubescens
(Del. & Pschw.) et vitripennis (Meigen) du genre Heringia Rondani (tribu des Pipizini). Concernant la
sous-famille des Syrphinae, il en est de même pour les femelles appartenant au sous-genre
Pandasyophtalmus Stuck. du genre Paragus Latreille, et pour les femelles des espèces bankowskae
Goeldlin, batava Goeldlin, chongjini Bankow., philantha (Meigen) et, dans une moindre mesure,
taeniata (Meigen) du groupe scripta du genre Sphaerophoria Le Pel. & Serville.
Régulièrement, des travaux de recherche en systématique et des révisions de genres (GOELDLIN DE
TIEFENAU, en préparation pour le genre Sphaerophoria, HURKMANS, 1993 et en préparation pour le
genre Merodon) et de tribus (MAIBACH et al., 1994a, 1994b pour les Chrysogasterini) apportent de
nombreux changements dans la nomenclature des Syrphides, ce qui nécessite une veille
bibliographique constante.
Nous sommes en droit de nous demander si le recours aux techniques très récentes d'investigation
systématique, à savoir celles concernant la biologie moléculaire (largement utilisées en mycologie
notamment), ne serait pas susceptible d'apporter une aide efficace dans l'étude de tous ces
problèmes d'identification des Syrphidae.
III - Biogéographie :
Les Syrphidae comprennent plus de 5000 espèces connues dans le monde, sont présents sur tous
les continents hormis l'Antarctique, et appartiennent à une des plus vastes familles de Diptères ; la
région paléarctique à elle seule compte près de 1600 espèces réparties en 120 genres (PECK, 1988).
La France, dont la faune des Syrphides est actuellement en cours de révision (SPEIGHT, 1993, 1994,
SPEIGHT et al., en préparation) totalise à l'heure actuelle environ 480 espèces (SPEIGHT, com. pers.),
réparties en 78 genres. Cette révision se base sur l'examen des collections du Muséum National
d'Histoire Naturelle de Paris, de même que de celles d'autres Muséums européens. Elle s'appuie
d'autre part, sur de récents inventaires régionaux activement effectués pour l'essentiel par des
syrphidologues de pays voisins : SPEIGHT (1984, 1988b et sans référence), DIRICKX & STEFFEN
(1992), CADOU (1993), BAUSENWEIN (1993), DIRICKX et al. (1996), DUSSAIX (en préparation), SARTHOU
(en préparation). Cette révision a permis de signaler jusqu'à maintenant 45 espèces nouvelles pour
la France ; depuis SEGUY (1961), la liste française s'est allongée de 170 espèces.
Le rythme d'allongement de cette liste au cours de la révision, soit de 1992 à aujourd'hui, est
révélatrice de la méconnaissance que nous avions de notre faune et de son importance : 436
espèces en janvier 1995(1) , 454 espèces en octobre 1995(1), 480 espèces en France en juin 1996(1)
(316 espèces mentionnées en 1961 par E. SEGUY).
Dans tous les pays bordant la Manche et la Mer du Nord, des listes d'espèces, au moins régionales,
ont été dressées récemment : Pays-Bas (BARENDREGT, 1982), Allemagne (BARKEMEYER, 1984, 1986,
CLAUSSEN, 1980, 1982, 1984, 1986), Grande-Bretagne (STUBBS & FALK, 1983) 252 espèces à l'heure
actuelle(2), Irlande (SPEIGHT, 1985) 170 espèces à l'heure actuelle(2), Danemark (TORP, 1984) environ
270 espèces à l'heure actuelle(2), Belgique (VERLINDEN & DECLEER, 1987 ; VERLINDEN, 1994) environ
313 espèces à l'heure actuelle(2).
MAIBACH et al. (1992) ont publié la liste faunistique des Syrphidae de Suisse (environ 415 espèces) et
SPEIGHT & LUCAS (1992) celle du Liechtenstein (187 espèces).
Depuis quelques années, la syrphidofaune ibérique fait l'objet d'études faunistiques et écologiques
nombreuses attestées par la série d'articles de MARCOS-GARCIA de 1981 à 1990 et de MARCOS-
(1) SPEIGHT, com. pers.
(2) SPEIGHT, com. pers., qui précise que de nombreuses mises à jour sont nécessaires, certaines en cours.

Synthèse bibliographique sur les Syrphidés – Thèse de Jean-Pierre SARTHOU 1996
4
GARCIA & CLAUSSEN (1989), MARCOS-GARCIA & LASKA (1983) et MARCOS-GARCIA & ROJO (1994) mais
aucune liste exhaustive d'espèces n'a encore été publiée.
Il apparaît que la syrphidofaune de France, pourtant prospectée tardivement et de façon peu
intensive, est la plus riche de toutes celles des pays européens et renferme plus de 50% des
espèces de la syrphidofaune européenne totale (SPEIGHT et al., en préparation). Ces recherches
faunistiques en Europe de l'Ouest font suite à d'importantes prospections et intègrent les nombreux
progrès en systématique de ces dernières années.
Parallèlement, des études biocénotiques portant sur la dynamique spatio-temporelle des
peuplements et la phénologie des espèces ont été menées à l'échelle d'un paysage (HÖVEMEYER,
1987 ; MARCOS-GARCIA, 1987 b ; OWEN & GILBERT, 1989 ; SSYMANK, 1991) ou plus largement de
provinces biogéographiques afin de comprendre la phylogénie et la structuration des peuplements
(HIPPA, 1978a, 1978b, 1990 ; MARCOS-GARCIA, 1987b).
IV - Morphologie :
A - Adultes :
Bien que la plupart de ses représentants adultes se reconnaissent immédiatement à courte distance,
la famille des Syrphidae se définit exclusivement par des caractéristiques alaires :
présence de deux nervures (MP1a, CuA1a), à l'extrémité apicale de l'aile, plus ou moins
parallèles au bord postérieur qui reste membraneux (ou bordé d'une sclérification punctiforme
chez les genres Episyrphus, Meliscaeva et chez Meligramma cincta) ; elles forment ensemble le
faux-bord postéro-apical.
présence d'une vena spuria (ou fausse nervure), simple pli longitudinal plus ou moins sclérifié
qui parcourt la plus grande partie des cellules r4 et r5 et qui traverse donc la transverse r-m
mais dont les deux extrémités (apicale et distale) sont libres, c'est-à-dire non rattachées aux
autres nervures. Il faut noter que cette vena spuria peut être chez de rares espèces, très peu
marquée à quasi absente (Eristalinus sepulchralis, Psilota anthracina, Syritta flaviventris).
Quant aux caractéristiques secondaires (arista implantée sur la base du troisième article
antennaire, absence de chète fort sur le thorax et plus généralement sur tout le corps, finesse des
pattes...), il y a toujours un nombre plus ou moins important d'exceptions.
L'habitus des Syrphidae adultes est en effet extrêmement varié : la longueur totale oscille entre 3,5
mm (Neoascia spp.) et plus de 24 mm (Milesia crabroniformis) ; le corps peut être large et robuste
(ex. : Eristalis tenax) mais aussi très mince et allongé (ex. : Baccha elongata).
La tête est généralement occupée en plus grande partie par les yeux composés d'ommatidies parfois
de taille variable entre les parties supérieure et inférieure de l'oeil (ex. : les mâles de Scaeva dignota
et de S. selenitica). Les yeux sont généralement dichoptiques chez les femelles et holoptiques chez
les mâles sauf exceptions comme pour les espèces Neoascia spp., Helophilus spp., Parhelophilus
spp., Anasimyia spp., Eristalinus sepulchralis... dont les mâles ont eux aussi des yeux dichoptiques
mais plus rapprochés que chez les femelles. Les antennes, toujours composées de trois articles,
sont implantées entre les yeux soit directement sur la capsule céphalique, soit sur une apophyse peu
développée (ex. : Pipiza spp.) ou plus rarement sur une apophyse assez développée (ex. : Psarus
sp.) à très développée (Ceriana spp.). Entre les yeux et au-dessus des antennes, se trouve le front,
totalement glabre ou plus ou moins recouvert d'une fine pubescence argentée appelée pruinosité et
dont la partie supérieure forme le vertex qui porte les ocelles, disposés en triangle équilatéral ou
isocèle. La face se trouve sous les antennes et possède généralement une protubérance centrale
plus ou moins saillante : le calus facial. L'orifice buccal est large pour contenir le proboscis rétractile
(non examiné pour la détermination des espèces) et est bordé par l'épistome qui constitue le rebord
inférieur de la face ; les genae, entre les yeux et l'ouverture buccale, et les zygomae, ourlets entre la
marge intérieure des yeux et la face, sont plus ou moins développés et ciliés de même que l'occiput,
rebord postérieur de la tête, derrière les yeux.

Synthèse bibliographique sur les Syrphidés – Thèse de Jean-Pierre SARTHOU 1996
5
Le thorax comprend antéro-dorsalement le mésonotum (avec les calus huméraux en position
antérieure et les calus postalaires en position postérieure), suivi du scutellum, dont la pilosité et la
coloration varient selon les espèces ; les côtés du thorax constituent les pleures formés de plusieurs
parties dont seules quelques-unes sont importantes dans la détermination des espèces : la partie
antérieure plane du mésopleure, le sternopleure et le métasternum (ciliés ou non).
Quant aux ailes, ce sont la disposition des nervures, donc la forme des cellules, la coloration, la
présence ou l'absence de microtriches dans certaines cellules ou sur l'alule, qui sont importantes à
considérer. Les cuillerons thoraciques ou squames, petites membranes reliant la partie basale de
l'aile au thorax, sont généralement sans longues soies jaunes (sauf chez le genre Syrphus).
Les pattes sont normalement simples (avec 5 articles aux tarses), mais les mâles, et parfois dans
une moindre mesure les femelles, de certaines espèces (ex. : Syritta pipiens, certains Brachypalpus,
Tropidia, Chalcosyrphus et Merodon) peuvent présenter un fémur postérieur renflé, muni ou non
d'épines. Les mâles de certains genres portent des apophyses spécifiques sur les coxae ou les
trochanters (ex. : Heringia spp.).
L'abdomen peut être de forme diverse, court et triangulaire (ex. : Merodon spp., Eristalinus spp.) ou
allongé et lancéiforme (ex. : Doros spp.) ; il est composé de trois ou quatre (parfois cinq) tergites bien
visibles, sauf exception (deux seulement chez Triglyphus primus). Les derniers segments sont
rudimentaires et portent (mm) ou protègent (ff) les genitalia, dont la structure chez les mâles est
assez complexe et variable entre genres et espèces, donc utile à l'identification de celles-ci.
L'abdomen (et parfois le thorax) est souvent marqué de taches ou bandes claires conférant ainsi aux
Syrphes "une allure de guêpe" ; d'autres espèces ont un corps entièrement noir leur donnant "l'allure
classique d'une mouche". Ainsi, le mimétisme est très répandu chez les Syrphidae et bien que la
ressemblance de certaines espèces avec des guêpes, des abeilles ou des bourdons puisse être
relative, il existe des cas de mimétisme quasi parfaits (Temnostoma spp., Brachypalpus spp.,
Criorhina pachymera, C. ranunculi). Le Syrphide Ceriana conopsoïdes est mimétique justement des
Conopidae (Diptères) des genres Conops et Physocephala.
Il est par ailleurs assez fréquent de noter des variations de coloration des ornementations
abdominales chez certaines espèces comme Episyrphus balteatus, Meliscaeva auricollis,
Sphaerophoria scripta, Eristalis arbustorum et Eristalis tenax par exemple. Il s'agit d'espèces
polyvoltines dont les générations de début du printemps et d'automne renferment une proportion
élevée d'insectes foncés n'ayant que peu de parties claires sur l'abdomen. Ceci suggère qu'en plus
d'un facteur d'ordre génétique (HEAL, 1989), intervienne un déterminisme d'ordre thermo-
physiologique : les températures basses agissent pendant le développement nymphal en favorisant
l'extension des taches sombres comme l'ont montré DUSEK & LASKA (1974), HEAL (1981) et
HOLLOWAY (1993).
Pour une description détaillée et complète de la morphologie externe des Syrphidae, nous renvoyons
au travail de SPEIGHT (1987).
B - Oeufs :
Tous les oeufs connus de Syrphides ont la même apparence : blanchâtres, de forme ovoïde parfois
arquée, allongés, avec une extrémité plus étroite que l'autre. Leur taille varie selon celle des espèces
(de 800 à 1300 µm environ) de façon proportionnelle généralement, bien que les oeufs de Syritta
spp. et Xylota spp. soient, par rapport à la taille des imagos, assez petits (GILBERT, 1986a). Le
chorion, sécrété par les cellules folliculaires et constitué de deux membranes microporeuses, assure
les fonctions de protection et de respiration en permettant à l'oeuf d'absorber l'oxygène de l'air et de
l'eau (KUZNETSOV, 1988b). Cette faculté adaptative est importante lorsque l'oeuf est entièrement
recouvert d'eau lors de rosées abondantes. La membrane externe du chorion, ou exochorion, a une
structure superficielle sculptée en réseaux ou en stries longitudinales. Plusieurs auteurs ont mis en
évidence le caractère spécifique de ces sculptures et ont réalisé des clés d'identification spécifique
des oeufs à partir de l'observation des chorions au microscope optique (KABOS, 1943 ;
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
1
/
27
100%