Ronéo

I) Electrophysiologie :
1) Introduction, les fonctions générales du système nerveux :
Dans une approche globale, on peut réduire la fonction du système nerveux à assurer la vie de
relation. C'est-à-dire, permettre l’élaboration de comportements adaptés et assurer
l’homéostasie interne.
En général, les comportements se traduisent par une activité motrice, de plus ils sont
déterminés par des facteurs externes et des paramètres internes. Par exemple s’il fait froid on
va s’habiller, ou si on a faim on va manger.
Schématiquement ce qui motive les comportements sont la survie, la reproduction et les
systèmes de récompense. De nos jours, survie et reproduction sont assurés et donc la
principale motivation va être représentée par les récompenses (l’obtention d’un diplôme, faire
du money cash,…).
L’homéostasie concerne l’équilibre du milieu interne. Le maintien de sa stabilité va dépendre
des paramètres internes et parfois des comportements.
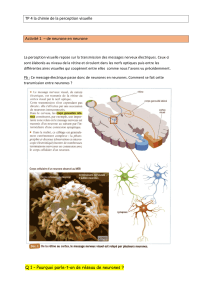
Comment fonctionne le système nerveux?
Grossièrement, voici un schéma qui résume son fonctionnement.
La stimulation se propage
par le SNP puis emprunte la
moelle : soit directement par
une voie rapide (avec peu de
synapse), soit une voie avec
de multiples étapes. Cette
dernière rend possible les
réflexes (vers une réponse
motrice ou une adaptation
du milieu interne).
En général, la stimulation
est de type sensitif et va
donc atteindre le cortex
sensitif. Celui-ci est relié au
cortex moteur et envoie ses
influx par la moelle (soit par
voie rapide ou multi
synaptique) pour engendrer
une réponse motrice ou une
adaptation du milieu
intérieur.
Vous trouverez ci-dessous un autre schéma avec plus de détails sur les acteurs et les réponses
obtenues, lors du fonctionnement du système nerveux.

2) le neurone (unité fonctionnelle du Système Neveux) :
a-Quelques chiffres:
Le corps humain contient environ 100 milliards de neurones (autant d’étoiles que dans la voie
lactée), cependant les cellules gliales sont 10 fois plus nombreuses.
Chaque neurone entre en contact avec 10 000 autres neurones par l’intermédiaire de synapse
(contrairement à ce que l’on voit dans les schémas simplifiés, où un neurone est en contact
avec un seul autre).Cet ensemble fonctionne en même temps avec des synapses qui sont soit
inhibitrice, soit excitatrice et par conséquent on obtient sur un neurone une réponse qui n’est
pas binaire. En outre c’est la somme des actions des 10 000 synapses qui détermine s’il y’a ou
pas un potentiel d’action.
Chaque jour environ 10 000 neurones disparaissent.
Le cerveau pèse entre 1.3 à 1.5 kg (soit 2% de la masse corporelle) et consomme 20% de
l’oxygène. L’énergie consommée est suffisante pour faire fonctionner une ampoule de 10
watts.
b-Caractéristiques du neurone:
C’est une cellule polarisée, c'est-à-dire que l’on peut mesurer une différence de potentiel entre
l’intérieur et l’extérieur de la membrane de -70mV. Notez que c’est cette différence qui
permet au neurone d’envoyer des messages électriques par des ondes de dépolarisation.
Ensuite, le neurone ne se multiplie pas sauf certaines régions, notamment, au niveau des zones
olfactives.
Une de ses autres caractéristiques est qu’il est spécialisé dans la signalisation et qu’il
communique au travers de synapses.
Enfin les neurones ont des formes variées adaptées à leurs fonctions mais il existe cependant
un schéma général (que l’on étudie dans la partie suivante).

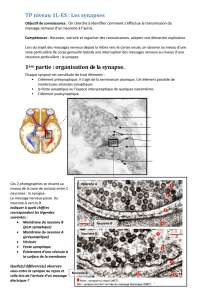
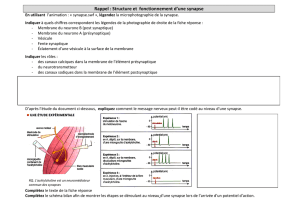
c-Morphologie fonctionnelle du neurone:
Dans chaque neurone on va trouver: -une zone réceptrice. Elle forme une couronne autour du
corps cellulaire et comprend les dendrites (ce sont des prolongements qui partent du corps
cellulaire et qui vont faire synapses avec les autres neurones).
Remarque: il existe aussi des synapses sur le corps cellulaire ou sur l’axone mais en générale
elles se font sur les dendrites.
-la zone intégratrice: c’est le corps cellulaire. Il va traiter
les signaux.
-la zone génératrice du potentiel d’action: c’est la zone
initiale de l’axone, appelée cône axonique ou segment initiale. C’est une zone spéciale très
riche en canaux ioniques.
-L’axone: c’est un câble qui va transporter le signal
nerveux jusqu’à la partie pré-synaptique de la synapse appelée « bouton terminal ».
-le bouton terminal : c’est la zone où sont stockées les
vésicules de Neurotransmetteur.
d-Propriétés de la membrane.
C’est une membrane faite d’une bicouche lipidique avec une zone externe hydrophile et une
interne hydrophobe.
Normalement cette membrane est imperméable aux ions et donc pour qu’il y ait échanges
ioniques, il faut que soient présents orifices à travers celle-ci. Ces orifices sont des canaux
aqueux entourées de sous unité protéique : soit les bon vieux canaux que l’on connaît. Ils
permettent le passage de solution ionique entre les milieux extra et intra cellulaire. Il faut

savoir qu’ils sont spécifiques vis à vis de certains ions, en raison de la taille du diamètre du
canal.
A présent nous allons voir les différents modes d’ouverture de ces canaux.
D’abord le plus simple, il peut être ouvert en permanence et donc provoquer une fuite ionique
selon le gradient de concentration (du plus vers le moins). Par exemple le canal K+, la sortie
du potassium va provoquer une modification de la polarité de la membrane.
Ensuite il existe le canal voltage dépendant, il ne s’ouvre qu’en cas de dépolarisation ce qui
permet la propagation de l’influx par ouverture, de proche en proche, de ces canaux le long de
l’axone.
Moreover, le canal lié à un ligand. C’est le récepteur spécifique de la partie post-synaptique
que l’on appelle aussi le « récepteur canal », il possède un site sur lequel vient se fixer le
ligand (le neurotransmetteur dans la synapse), cette fixation va ouvrir le canal et permettre les
échanges ioniques.
Enfin, le canal dont l’ouverture dépend d’un signal intracellulaire.
3) Electrophysiologie du neurone.
a-cellule électriquement polarisée :
Comme on l’a dit, le neurone est une cellule électriquement polarisée: il y a donc une
différence de potentiel de -50 à -70 mV entre l’extérieur et l’intérieur de celle-ci qui est due à
une différence de concentration ionique ainsi qu’aux anions organiques (protéines intra
cellulaire).
Cette différence de concentration ionique, de part et d’autre de la membrane, est liée à un
processus actif, grâce à des pompes ioniques qui nécessite de l’énergie (ATP).
Par conséquent si la cellule meurt, il y a arrêt des mécanismes de maintien et l’équilibre des
ions se fait selon une diffusion à travers la membrane.
Voici donc un schéma expliquant la différence de potentiel avec le sodium et le potassium.
La tendance naturelle du sodium
est une diffusion de l’extérieur
vers l’intérieur.
Pour maintenir le gradient il y à la
pompe Na/K ATPase.
Le potassium a un gradient de
concentration inverse, il a donc
une tendance normale de sortie.
D’autre part, on a vu qu’il y avait
des canaux de fuite au potassium
ouverts en permanence. Donc le
K+ diffuse sans arrêt de l’intérieur
vers l’extérieur.

b) Evénements électriques au niveau du neurone:
Au niveau du neurone, tout part d’un potentiel local qui porte plusieurs noms: gradué ou
récepteur (selon l’endroit où on le trouve).
Lorsque ce potentiel local se somme et atteint un seuil il va y avoir production d’un potentiel
d’action qui va parcourir l’axone et arriver au niveau de la synapse. Ensuite il y a libération de
Neurotransmetteurs qui vont aller se fixer sur des récepteurs post-synaptique afin de générer,
soit un Potentiel Post Synaptique Excitateur (PPSE), ou un Potentiel Post Synaptique
Inhibiteur (PPSI).
Enfin, il y aura des phénomènes d’intégration au niveau de cette synapse.
Voici un schéma pour mettre en situation ces notions.
PPSI (qui sont des potentiels locaux au niveau d’une synapse).
3) Si ces PPSE sont suffisants ils vont s’additionner et si la somme atteint un seuil il va y
avoir émission d’un potentiel d’action à partir du cône axonique.
4) Le potentiel d’action se propage et arrive au niveau du muscle provoquant un potentiel de
plaque (qui est lui aussi un potentiel local).
5) si le potentiel de plaque est suffisant, il va produire un potentiel d’action musculaire.
Donc à l’origine de tous les potentiels d’action, qu’il soit sensitif ou moteur, etc…, il y a des
potentiel locaux appelés potentiel de récepteur (si c’est au niveau d’un récepteur), PPSE ou
PPSI (si c’est au niveau d’une synapse) et potentiel de plaque (au niveau d’une plaque neuro
musculaire).
(Remarque : personnellement je suis gêné d’intégrer les PPSI quand on sait qu’ils empêchent
l’activation des neurones. Mais bon je suis pas physiologiste.).
Ici c’est l’exemple d’un
récepteur en périphérie,
on a donc un petit
potentiel local dit de
« récepteur ».
1) Lorsque l’on stimule
la peau, on va
provoquer des
potentiels au niveau du
récepteur qui vont
s’additionner.
2) Lorsqu’ils
atteignent un seuil, ils
vont déterminer la
production d’un
potentiel d’action qui
se propage le long de
l’axone et va arriver au
niveau de la synapse
3) cela a pour
conséquence la
production de PPSE ou
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%