Corrigé Bac blanc de SVT

Indicateur de correction partie I – bac blanc – avril 2006
Introduction
Le VIH (virus de l'immunodéficience humaine) est un virus qui a pour hôte l'organisme humain. Comme tout virus, il ne peut réaliser son cycle de
développement qu'à l'intérieur de cellules, en l'occurrence les cellules du système immunitaire comme les lymphocytes T4.
Entre l'entrée du virus et l'apparition de la maladie, le SIDA, a lieu une évolution au cours de laquelle le système immunitaire tente d'éliminer le
virus, mais souvent sans succès.
Après avoir expliqué les modalités de l’infection virale, nous montrerons comment, en l’absence de traitement efficace, cette infection conduit à
l’apparition du SIDA.
I. Le cycle de développement du VIH
1. Un rétrovirus à ARN
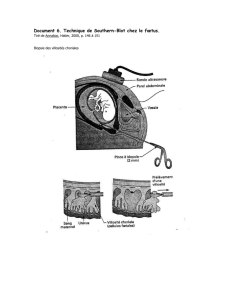
Le VIH, pendant la phase extracellulaire de son cycle, est une particule sphérique délimitée par une enveloppe lipidique au niveau de laquelle sont
fixées, vers l'extérieur, des protéines appelées gp 120.
À l'intérieur, une autre enveloppe, de nature protéique, la capside, renferme deux molécules d'ARN et quelques enzymes nécessaires au cycle viral.
C'est cet ARN qui porte l'information génétique du virus et qui permet de le classer dans la catégorie des rétrovirus.
Structure du VIH.
2. Le cycle de développement du VIH
Le VIH est incapable de se multiplier à l’extérieur des cellules de l'organisme. Cela s'explique par l'absence d'organites et d'enzymes nécessaires.
Pour se développer, il doit pénétrer dans les cellules de l'organisme comme les lymphocytes T4. Cela commence par la fixation du virus (étape
sur le schéma), par ses protéines gp120, à des récepteurs présents principalement sur la membrane plasmique des lymphocytes T4, (LT4), et de
forme complémentaire, les molécules CD4.
C'est ce qui explique que ces cellules sont une cible pour le VIH. Cette fixation provoque la fusion de l'enveloppe virale avec la membrane
plasmique des LT4. La capside est injectée dans le LT4 et l'ARN viral libéré dans le cytoplasme.
Intervient alors une enzyme virale fondamentale, la transcriptase inverse. Elle réalise une transcription à l'envers, c'est-à-dire la synthèse d'une
molécule d'ADN double brin à partir d'un ARN simple brin. Une autre enzyme, l'intégrase intègre alors l'ADN viral dans l'ADN cellulaire.
À ce stade, le virus ne subsiste plus dans la cellule que sous la forme d'une molécule d'ADN.
Vue d'ensemble de l'infection d'une cellule cible par le VIH.
Cet ADN va ensuite être transcrit en ARN, lui-même traduit en protéines virales. Cette activité n'est possible que par l'intervention
d'enzymes et d'organites cellulaires, comme les ribosomes, grâce à l'énergie fournie par la cellule. C’est ce qui explique que le virus ne peut pas se
reproduire à l'extérieur de la cellule.
Enfin, les protéines virales vont subir des transformations puis être assemblées avec l'ARN. La membrane plasmique du LT4 bourgeonne,
englobant avec elle des virus qui sont alors libérés hors de la cellule.
Le cycle pourra alors recommencer avec de nouveaux LT4.

II. La primo infection (entrée du VIH dans l'organisme et la phase asymptomatique
L'entrée du virus dans l'organisme s'accompagne de la mise en route du cycle, sans pour autant déclencher la maladie. Le virus se multiplie
activement pendant quelques mois. C'est la phase aiguë.
Des LT4 reconnaissent le virus, sont activés et sécrètent des interleukines permettant la prolifération et la différenciation de lymphocytes B et de
lymphocytes T cytotoxiques dirigés contre le VIH et les cellules infectées.
Cette réponse entraîne une diminution notable du nombre de VIH.
La présence du VIH dans l'organisme peut être détectée par la présence d'anticorps anti-VIH dans le sérum de l'individu. Celui-ci est alors qualifié
de séropositif pour le VIH.
Pendant cette phase, qualifiée d'asymptomatique car l'individu n'est pas malade, le nombre de LT4 baisse, ceux-ci étant détruits directement ou
indirectement par le virus. En effet, le virus échappe progressivement au système immunitaire, notamment par son haut pouvoir de mutation.
Néanmoins, le nombre de LT4 reste suffisant pour participer aux réactions immunitaires contre les micro-organismes pénétrant dans le corps.
Cette phase a une durée très variable. On connaît même des individus résistants à la maladie, restant asymptomatiques alors qu'ils ont été infectés
depuis plus de deux décennies.
Néanmoins, pendant cette phase, le sujet reste contagieux, le virus pouvant être transmis à l'intérieur des LT4 par les liquides qui les contiennent,
sang, sperme et sécrétions génitales notamment.
III. Le développement de la maladie ou phase symptomatique
L'apparition de la maladie est déclenchée par la chute du nombre de LT4. En dessous d'une certaine quantité de LT4, l'organisme n'est plus
capable de se défendre contre les divers micro-organismes présents dans son environnement. La diminution de LT4 ne permet plus la coordination
habituelle de la réponse immunitaire. En effet, la sécrétion d'interleukines baisse et le nombre de lymphocytes B et T différenciés diminue.
Les trois étapes du processus infectieux après contamination par le VIH,
en l'absence de traitement.
Ainsi, avec moins de molécules et de cellules effectrices, le système immunitaire n'est plus capable de neutraliser les micro-organismes comme il le
fait en temps normal. II subit donc une succession d'infections qualifiées d'opportunistes car ces micro-organismes profitent de l'opportunité de
cette immunodéficience de l'organisme ne peut plus lutter contre ces infections et finit par en mourir.
Ce n'est donc pas le VIH qui entraîne directement la mort, même s'il en est la cause première.
Bilan
Le VIH a pour cible principale les LT4, véritables pivots du système immunitaire. De ce fait, il conduit généralement et en l’absence de traitement
efficace, à une immunodéficience généralisée aux conséquences mortelles.
Les progrès dans la connaissance du cycle viral ont permis de développer des molécules qui bloquent celui-ci. Aujourd'hui, on travaille sur la mise
au point d'un vaccin qui stimulerait Ie système immunitaire dans sa lutte contre le VIH.

Barème 2a 04/2006
barème
Saisie des données
Interprétations
barème
/0.5
/1
Après recombinaison chromosomique par crossing over
au cours de la méiose chez un individu masculin
:
- une chromatide de X porte anormalement le gène
Sry
- une seule des deux chromatides de Y porte le gène
Sry
-Si présence du gène sry apparition du phénotype
masculin
- mais si absence du gène Sry apparition du
phénotype féminin
/0.5
/0.5
Mise en relation :
XX caryotype normalement féminin, la présence du gène Sry le chromosome X d’origine paternelle, est
responsable de l’apparition du phénotype masculin.
XY caryotype normalement masculin , l’absence du gène Sry sur le chromosome Y détermine l’apparition d’un
phénotype féminin.
/1
Pas d’erreur fondamentale, la démarche scientifique est correcte (non confuse)
/0.5
Partie 2b obligatoire : stabilité et variabilité des génomes et évolution
barème
Saisie des données
Interprétation
Connaissances conclusions
barème
0.25
0.25
0.5
0.5
Document 1 : transmission simultanée de deux caractères :
deux hypothèse : gènes indépendants ou liés
Le codage utilisé pour les allèles est défini
croisement 1
parents homozygotes, uniformité F1
Croisement 2 :
Majorité de phénotypes parentaux, Minorité de
phénotypes recombinés
phénotypes, convention d’écriture formulée [ ]
gris>noir, soies lisses> crochues
F1 hétérozygote
Croisement test cité
signification des phénotypes issus du test cross, précisée (les
différents types de gamètes produits par l’hybride)
Si 2 gènes indépendants 4 combinaisons alléliques
équiprobables
Réfutation hypothèse gènes indépendants
ou gènes liés , 4 phénotypes non équiprobables
confirmation gènes liés
0.25
0.25
0.5
0.5
0.5
0.25
0.25
0.25
Document 2 : prophase de méiose chez la femelle de F1
figure en prophase de méiose chez la femelle F1,
chiasma entre chromosomes homologues
crossing over entre deux chromosomes homologues chiasma =
point d’échange entre deux chromatides
Deux combinaisons alléliques minoritaires en F2,
expliquées par l’échange entre les homologues,
de portions de chromatide portant les allèles
d’un des deux gènes. (ici ou ailleurs)
0.25
0.25
Mise en relation :
Interprétation des résultats
Bonus tableau du croisement test exact - - - - - - - - - -- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
Schéma exact, du crossing over avec allèles situés au dessus et au dessous du chiasma (ou avant après) ,
schéma acceptable dans sa forme
génotype de la F1 exact, conventions d’écriture respectées ( / ) ou ( // )
+ 0.5
0.5
0.25
0.5

2B spécialité : Corrigé indicatif, non rédigé
Introduction : reformuler la problématique, ne pas définir la théorie de l’hérédité par mélange qui doit l’être à partir de l’exploitation du document1
LA THEORIE DE L’HEREDITE PAR MELANGE : Document 1 :
Largement admise à l’époque de Mendel, théorie selon laquelle les caractères des parents fusionnent dans la descendance sans possibilité de
séparation ultérieure. Les descendants ont des caractères intermédiaires entre ceux des parents
APPORTS DES TRAVAUX DE MENDEL : Document 2 :
Expliquer brièvement le principe des travaux.
(Observations): Les données essentielles doivent être traduites de façon synthétique, afin de ne pas noyer le devoir, mais précise*:
- Issus du croisement de deux lignées pures (obtenues par autofécondations successives, un seul caractère stable), les individus de
F1sont tous identiques (phénotype uniforme) ne présente qu’un seul des deux caractères parentaux (caractère lisse).
- en F2 l’autre caractère (phénotype ridé) réapparaît. En F3, le caractère ridé ne donne que des caractères ridés par autofécondation, il
réapparaît donc dans toute sa pureté (termes de Mendel)
(Mise en relation des données pour interprétation):
- Le caractère qui réapparaît en F2 (ou en F3) est inchangé comme le montrent les résultats de l’autofécondation. Mendel explique qu’il était présent
chez les hybrides de F1(F2) mais ne se manifestait pas. Il formule la notion de caractère récessif. Le caractère qui est visible chez les hybrides, est
le caractère dominant.
- Le phénotype de la F1 n’est pas intermédiaire entre ceux des parents, la présence simultanée chez les hybrides (de 1ère ou de 2ème
génération) du caractère récessif et du caractère dominant, la réapparition du caractère récessif intact, indiquent que :
Il n’y a jamais fusion des caractères parentaux chez l’hybride,
Les deux caractères parentaux présents chez l’hybride (loi uniformité des hybrides de F1) se disjoignent lors de la formation des
gamètes qui ne contiennent qu’une version alternative d’un caractère donné (loi de pureté des gamètes)
Donc Mendel élimine le concept d’hérédité par mélange (doc 1).
Ce qui est transmis est un trait du caractère – une particule- qui est le support du caractère.
Donc Mendel formule le concept d’hérédité particulaire
* (A condition d’interpréter l’observation qui suit en mettant en évidence le caractère statistique des études de Mendel, on peut citer aussi :
- Les proportions obtenues pour les autofécondations des hybrides en F2 (F3) sont toujours ¾ -caractère présent en F1- et ¼ -caractère réapparu
en F2. Ainsi Mendel établit des résultats statistiques, reproductibles )
Remarque1 : aucune loi n’est attachée à ce dernier résultat
Remarque 2 : une analyse génétique n’est pas demandée ici, il faut donc raisonner à la façon de Mendel
Bilan. Les apports de Mendel : réfutation de la théorie d’hérédité par mélange, formulation d’un concept heuristique (ouvre la recherche scientifique
vers d’autres découvertes) le concept de l’hérédité particulaire est proche du concept de gène qui n’existe pas encore. Apr7s 10 années de travaux
minutieux, Mendel pose les bases théoriques de la génétique mais l’importance de ses travaux passe inaperçue pendant 40 ans
Ne pas parler de révolution de la génétique, celle –ci i n’existe pas avant Mendel. Ne pas exagérer l’importance des ses travaux au point d’en faire
dépendre les biotechnologies… Ne parler que de ce dont on est certain, une grosse bêtise nuit plus sûrement au devoir que des connaissances
réduites mais exposées clairement, une affirmation trop vague met en évidence la pauvreté des connaissances.
2B spécialité : Corrigé indicatif, non rédigé
Introduction : reformuler la problématique, ne pas définir la théorie de l’hérédité par mélange qui doit l’être à partir de l’exploitation du document1
LA THEORIE DE L’HEREDITE PAR MELANGE : Document 1 :
Largement admise à l’époque de Mendel, théorie selon laquelle les caractères des parents fusionnent dans la descendance sans possibilité de
séparation ultérieure. Les descendants ont des caractères intermédiaires entre ceux des parents
APPORTS DES TRAVAUX DE MENDEL : Document 2 :
Expliquer brièvement le principe des travaux.
(Observations): Les données essentielles doivent être traduites de façon synthétique, afin de ne pas noyer le devoir, mais précise*:
- Issus du croisement de deux lignées pures (obtenues par autofécondations successives, un seul caractère stable), les individus de
F1sont tous identiques (phénotype uniforme) ne présente qu’un seul des deux caractères parentaux (caractère lisse).
- en F2 l’autre caractère (phénotype ridé) réapparaît. En F3, le caractère ridé ne donne que des caractères ridés par autofécondation, il
réapparaît donc dans toute sa pureté (termes de Mendel)
(Mise en relation des données pour interprétation):
- Le caractère qui réapparaît en F2 (ou en F3) est inchangé comme le montrent les résultats de l’autofécondation. Mendel explique qu’il était présent
chez les hybrides de F1(F2) mais ne se manifestait pas. Il formule la notion de caractère récessif. Le caractère qui est visible chez les hybrides, est
le caractère dominant.
- Le phénotype de la F1 n’est pas intermédiaire entre ceux des parents, la présence simultanée chez les hybrides (de 1ère ou de 2ème
génération) du caractère récessif et du caractère dominant, la réapparition du caractère récessif intact, indiquent que :
Il n’y a jamais fusion des caractères parentaux chez l’hybride,

Les deux caractères parentaux présents chez l’hybride (loi uniformité des hybrides de F1) se disjoignent lors de la formation des
gamètes qui ne contiennent qu’une version alternative d’un caractère donné (loi de pureté des gamètes)
Donc Mendel élimine le concept d’hérédité par mélange (doc 1).
Ce qui est transmis est un trait du caractère – une particule- qui est le support du caractère.
Donc Mendel formule le concept d’hérédité particulaire
* (A condition d’interpréter l’observation qui suit en mettant en évidence le caractère statistique des études de Mendel, on peut citer aussi :
- Les proportions obtenues pour les autofécondations des hybrides en F2 (F3) sont toujours ¾ -caractère présent en F1- et ¼ -caractère réapparu
en F2. Ainsi Mendel établit des résultats statistiques, reproductibles )
Remarque1 : aucune loi n’est attachée à ce dernier résultat
Remarque 2 : une analyse génétique n’est pas demandée ici, il faut donc raisonner à la façon de Mendel
Bilan. Les apports de Mendel : réfutation de la théorie d’hérédité par mélange, formulation d’un concept heuristique (ouvre la recherche scientifique
vers d’autres découvertes) le concept de l’hérédité particulaire est proche du concept de gène qui n’existe pas encore. Apr7s 10 années de travaux
minutieux, Mendel pose les bases théoriques de la génétique mais l’importance de ses travaux passe inaperçue pendant 40 ans
Ne pas parler de révolution de la génétique, celle –ci i n’existe pas avant Mendel. Ne pas exagérer l’importance des ses travaux au point d’en faire
dépendre les biotechnologies… Ne parler que de ce dont on est certain, une grosse bêtise nuit plus sûrement au devoir que des connaissances
réduites mais exposées clairement, une affirmation trop vague met en évidence la pauvreté des connaissances.
1
/
5
100%