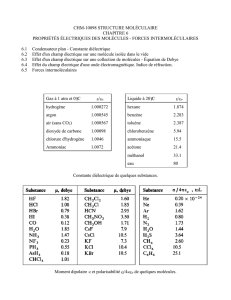
Immunité spécifique Elle fait intervenir des lymphocytes qui sont

Immunité spécifique
Elle fait intervenir des lymphocytes qui sont issus des organes lymphoïdes primaires (mœlle
osseuse et thymus pour les T). Les lymphocytes vont reconnaître l'antigène par des récepteurs
spécifiques (on dit que chaque lymphocyte est programmé pour reconnaître un antigène donné) et
vont s'activer pour donner 2 catégories de cellules:
- effecteurs qui vont éliminer l'antigène.
- lymphocytes mémoire.
Les antigènes sont apportés par les cellules dendritiques dans les organes lymphoïdes secondaires.
Présentation et reconnaissance de l'antigène
Le système immunitaire utilise 3 systèmes:
- molécules du CMH (complexe majeur d'histocompatibilité) pour la présentation de
l'antigène.
- TCR (T cell receptor)
- BCR (B cell receptor)
Le complexe majeur d'histocompatibilité
I) Historique
En 1938, P. Gorer réalisait des greffes de peau chez des souris de souches différentes et constatait
qu'elles étaient toujours rejetées. Il s'agissait d'un problème existant entre donneur et receveur. Il a
ensuite observé que ce rejet était sous la dépendance du système immunitaire.
Il a fallu attendre les travaux de G. Snell, qui a identifié chez la souris un système qu'il a appelé le
système H2 (= CMH de la souris).
Chez l'Homme, J. Dausset a découvert, sur les leucocytes en 1958, les molécules qu'il appelle
''antigènes d'histocompatibilité'', et les regroupe dans un système qu'il appelle ''système HLA''. J.
Dausset les a appelé antigènes car il les a identifié grâce à des anticorps.
Il a fallu attendre les travaux de Doherty et Zurkernegel pour comprendre que ces molécules
jouaient un rôle physiologique fondamental: elles assurent une communication indispensable à la
réponse immunitaire, car elles assurent la présentation de l'antigène aux lymphocytes T.
Les travaux de P. Bjorkman ont permis de montrer comment ces molécules pouvaient assurer la
présentation de l'antigène.

HLA = Human Leucocyte Antigen
Ces molécules ont 2 noms chez l'Homme:
- Antigène d'histocompatibilité.
-
Une molécule du CMH est un antigène pour un individu qui ne la possède pas.
II) Définition du CMH
Le complexe majeur d'histocompatibilité est un ensemble de gènes localisés sur le bras courts du
chromosome 6. Il y a environ 200 gènes qui représentent environ 1/1000ème du génome humain.
C'est un système multigénique, multiallélique, et d'expression co-dominante. Parmi tous ces
gènes du CMH, il y en a qui codent des molécules qui vont présenter l'antigène aux lymphocytes T
et ces gènes constituent le système HLA.
III) Structure
Les molécules du système HLA sont des protéines transmembranaires qui appartiennent à la super-
famille des immunoglobulines car elles présentent un domaine immunoglobulin-like (environ 90
acides aminés) stabilisé par un pont disulfure.

Les molécules de classe I du CMH sont constituées d'une chaîne lourde et c'est à ce niveau là qu'on
retrouve la variabilité d'un individu à un autre. Cette chaîne lourde est constituée de 3 domaines α1,
α2 et α3, d'une partie transmembranaire et d'une courte partie cytoplasmique.
La chaîne lourde est associée à une chaîne légère qui est totalement invariante: β2-microglobuline.
Cette dernière n'est pas associée de manière covalente et est codée par un gène qui n'est pas dans le
CMH.
La molécule de classe II du CMH est formée de 2 chaînes:
- une chaîne α.
- une chaîne β.
Chaque chaîne a 2 domaines extracellulaires:
-α1 et α2
- β1 et β2
Chaque chaîne a une partie transmembranaire et une partie cytoplasmique très courte.

Les domaines les plus extracellulaires (α1 et α2, α1 et β1) vont adopter une configuration particulière.
Ils constituent une sorte de plateforme qui ménage une fente (25 angströms de long et 10 angströms
de large) où sera présenté le peptide.
La fixation du peptide est différente dans les 2 fentes.
- Pour la classe I, le peptide a moins de 9 acides aminés et il s'insère complètement dans la
cavité. Il envoie des acides aminés dans des poches spécifiques de l'allèle.
- Pour la classe II, le peptide peut faire plus de 9 acides aminés et il envoie aussi des acides
aminés dans des poches spécifiques de l'allèle.
Les molécules du CMH présentent un grand polymorphisme: d'un allèle à un autre, on a des acides
aminés qui changent au niveau de la cavité.
Une molécule de classe I ou II ne peut fixer qu'un peptide à la fois, mais elle peut fixer de très
nombreux peptides différents (on pense qu'elle peut fixer jusqu'à 1000 peptides différents). Comme
on ne dispose que d'un certain nombre d'allèles sur nos cellules, on ne peut pas tout présenter. Il y a
CMH II
CMH I

donc un tri cellulaire, en fonction des allèles qu'on a, ce qui explique également pourquoi tout le
monde ne répond pas de la même manière à une infection. Avec ce système, on n'a qu'une vue
partielle du Soi et du Non soi.
IV) Distribution tissulaire de ces molécules
Les molécules de classe I conventionnelles (HLA-A, HLA-B, HLA-C) ont une distribution
extrêmement ubiquiste. Dans le sang, toutes les cellules en possèdent à leur surface, sauf les
globules rouges. On les trouve partout sauf les hépatocytes, les cellules et les vaisseaux au niveau
central, le pancréas et le syncytiotrophoblaste.
Chaque fois qu'une cellule sera modifiée par un virus, par une transformation tumorale, les
molécules de classe I vont pouvoir avertir les lymphocytes T.
Les molécules de classe II ont une expression beaucoup plus restreinte. Dans le sang, on les trouve
uniquement sur 3 catégories de cellules qui présentent l'antigène aux lymphocytes T:
- lymphocytes B
- cellules dendritiques
- monocytes.
Dans les tissus, on les trouve au niveau des cellules endothéliales des capillaires et au niveau de
certaines cellules épithéliales de l'intestin grêle.
D'autres cellules peuvent présenter des molécules de classe II mais uniquement après stimulation.
V) Organisation génomique
On distingue 3 régions génétiques:
- région de classe I qui comporte des gènes qui codent pour les molécules HLA-A, HLA-B,
HLA-C, et les molécules HLA-E, HLA-F et HLA-G. On les appelle les molécules de classe
B car elles sont non conventionnelles (E → G)
- région de classe III qui comporte des gènes qui ne sont pas en relation directe avec le
système HLA. Il y a des gènes qui codent pour le complément (C2), pour des cytokines
(TNF), …
- région de classe II qui comprend des gènes qui codent pour les molécules de classe II du
CMH. Il y aura 2 gènes (A et B) pour chaque molécule car elles sont constituées de 2
chaînes. On appelle ces molécules HLA-DP, HLA-DQ et HLA-DR. Il y a d'autres gènes qui
codent des molécules qui interviennent dans le traitement et le transport des peptides (LMP
et TAP), et des gènes qui codent pour des molécules qui permettent le chargement des
peptides (DM, DO).
Pour tous ces gènes, il existe de très nombreux allèles (plus de 1000) → polymorphisme.
Exemples: pour HLA-B, il existe plus de 617 allèles.
pour HLA-A, il existe plus de 338 allèles.
 6
7
8
9
6
7
8
9
1
/
9
100%