La Première semaine de développement

La Première semaine de développement
1. Introduction
Les 2 premières semaines du développement de l’embryon débutent par la fécondation et finissent
par l’implantation ou nidation de l’œuf ou zygote dans l’endomètre. Cela correspond à la phase
sécrétoire, c'est-à-dire la 2ème phase du cycle menstruel.
Au cours de la première semaine de développement (SD1), l’œuf est libre dans les voies génitales
féminines, c’est la période de vie libre ou période pré-implantatoire. L’œuf se déplace dans la trompe
utérine en direction de la cavité utérine, on parle de migration tubaire. La mère ne présente aucun
signe clinique ou biologique de grossesse.
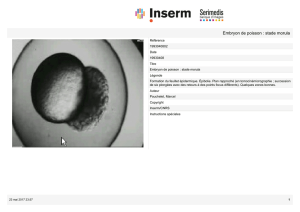
2. La segmentation de la Morula
L’œuf fécondé se divise très rapidement par mitose. La première division de segmentation va aboutir
à deux blastomères ou cellules-filles de taille identique.
Pendant les mitoses de segmentation suivantes, le nombre de blastomère augmente mais la taille du
zygote n’augmente pas (pas augmentation du volume). Les blastomères deviennent alors de plus en
plus petits, ils prennent une forme sphérique (ovoïde) et leur surface est couverte de microvillosités.
La segmentation permet la formation d’un massif cellulaire sphérique appelée morula dont les
cellules sont juxtaposées et indépendantes les unes des autres (échappement de la cellule si absence
de zone pellucide).
Les premières divisions sont sans activité de transcription. Les protéines sont traduites grâce aux
ARNm maternels. Les premières divisions utilisent donc les transcrits maternels. Le zygote fabriquera
ses propres protéines seulement après épuisement de ces transcrits (2-3ème jour de développement).
La période de clivage dure 4 jours. Pendant cette période, les blastomères sont totipotents, ils
expriment de façon équivalente la totalité du programme génétique. C’est lié à l’expression de oct3.

Les blastomère ont a ce stade une grande capacité de régulation : ils peuvent se remplacer les uns les
autres et chaque blastomère peut donner un zygote à part entière.
La totipotence des blastomères est perdue lors de la compaction.
3. La Compaction de la Morula
A ce stade, la morula comporte une vingtaine de blastomères. Intervient une activité génétique
différentielle entre les blastomères et l’acquisition d’une polarité. Elle fait intervenir les protéines de
la famille des CAM (Cell Adhesion Molecules) qui sont des protéines membranaires intervenant dans
la forme de la cellule ainsi que son adhérence intercellulaire.
Avant la compaction, les E-Cadhérines sont uniformément réparties sur la cellule. Lors de la
compaction, elles se regroupent aux points de contact intercellulaires. La compaction est déclenchée
par les E-Cadhérines.
Il y a aussi besoin de Ca++ qui permet la formation de liaison intercellulaires spécialisées qui relient
les blastomères entre eux.
On va avoir deux types de blastomères :
- Les blastomères périphériques qui vont subir la compaction et se polariser. Ils deviennent
plus petits et s’aplatissent. Ces cellules vont former le trophoblaste.
- Les blastomères centraux qui ne se polarisent pas et ne subissent pas la compaction. Ils
gardent leur forme sphérique et deviennent plus volumineux.
Le destin des blastomères est déterminé par leur position au sein de la Morula.

4. La Cavitation de la Morula
A ce stade, il y a une trentaine de blastomères au sein de la Morual.
Elle se déroule en plusieurs étapes :
- L’activité des pompes N/K ATPase crée un flux liquidien (flux de Na et courant osmotique).
- Le flux liquidien ne peut pas sortir de la morula (étanchéité au niveau du trophoblaste), il se
crée des lacunes intercellulaires dans la morula
- Les lacunes fusionnent et donnent une cavité appelée blastocèle
- La morula devient une sphère creuse que l’on appelle alors blastocyste ou blastula.
- La formation du blastocèle repousse la masse cellulaire interne à un pôle du blastocyste qui
est appelée bouton embryonnaire.
5. L’Inactivation d’un des deux chromosomes X ou lyonisation
Un individu est de sexe féminin dès qu’il a 2 chr X.
Bouton embryonnaire
Blastocèle
Trophoblaste

Au cours de la segmentation, un des deux chr X est compacté est s’inactive sous forme
d’hétérochromatine. Cet évènement est appelée lyonisation, il est à l’origine de l’apparission du
Corpuscule de Barr que l’on peut voir sur les cellules qui comportent au moins 2 chr X. Cette
inactivation se fait au hasard soit sur le chr X paternel ou soit sur le chr X maternel.
Ce n’est pas la totalité des gènes du Chr X qui sont inactivés. L’inactivation dépend du gène Xist. Elle
est irréversible pour les cellules somatiques mais réversible pour les cellules germinales avant leur
méiose.
6. La Migration Tubaire
Pendant la segmentation, le zygote migre du lieu de fécondation vers la cavité utérine. Cette
migration dépend de plusieurs facteurs : les contractions péristaltiques de la muqueuse tubaire, le
nombres et la qualités des cils vibratiles ainsi que de l’abondance du flux de liquide.
La migration dépend de 2 hormones :
- Les oestrogènes : ↗ sécrétion de liquide tubaire, ↗ la ciliogenèse (mouvement des cils)
- La progestérone : ↘ l’œdème de la paroie de l’isthme tubaire, ↗ le diamètre tubaire, ↗ la
contractibilité musculaire
Quand le zygote arrive dans la cavité utérine, il « divague » durant 2 jours. Sous l’effet de la
progestérone, le liquide utérin se charge en produit de sécrétion. Ceci est à l’origine de l’évolution du
zygote en blastocyste.
1
/
4
100%