CHAPITRE V

CHAPITRE V
DELIMITATION DE L'EMBRYON
ET DE SES ANNEXES
5.1. LA DELIMITATION DE L'EMBRYON
Jusqu'au stade trilaminaire, l'ébauche embryonnaire reste plate, étalée à la surface du
blastocyste. Sa face ventrale forme le plafond de la cavité vitelline et sa face dorsale répond à
la cavité amniotique chez les espèces à implantation précoce (primates, rongeurs,...) ou baigne
dans le milieu intra-utérin chez les espèces à implantation différée (ruminants, équins,
carnivores,...).
La gastrulation, c. à d. l'apparition de la ligne primitive, la détermination des trois axes et la
mise en place du mésoderme, se poursuit par la délimitation de l'embryon. Ce dernier
s'individualise en refermant sa face ventrale pour ne plus communiquer avec ses annexes que
par son cordon ombilical. Cette fermeture ventrale est la conséquence de la mise en place de
4 sillons marginaux au niveau desquels l'ectoderme se rabat vers la face ventrale de l'embryon
en refoulant devant lui les feuillets sous-jacents. Ces sillons marginaux deviennent de
véritables replis qui se rejoignent en région ombilicale. L'embryon "se replie" autour de son
cordon et se "recouvre de son amnios".
A) LE PLI CEPHALIQUE
Il procède d'une croissance plus rapide du feuillet ectodermique en région céphalique.
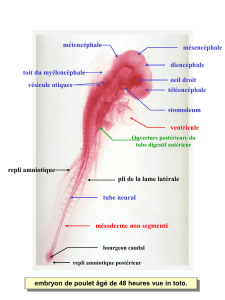
Ce pli adopte la forme générale d'un fer à cheval au centre duquel s'individualise le futur SNC
(figure 5.1). En vue longitudinale, le pli céphalique délimite la future cavité buccale au niveau
de la poche pharyngienne et se prolonge vers le pôle caudal en repoussant le massif
mésodermique où se forme le cœur (figure 5.1). Il forme de fait la partie antérieure du tube
digestif ou proentéron.
Le bord supérieur du pli recouvre progressivement l'embryon pour rejoindre le bord
correspondant des deux plis latéraux et du pli caudal. Il porte le nom de pli amniotique
crânial (figure 5.1).

B) LE PLI CAUDAL
Il est moins accusé que le pli céphalique. Il correspond également à une croissance
différentielle de l'ectoderme par rapport aux tissus sous-jacents. Il repousse ver le pôle crânial
la membrane cloacale et forme la partie postérieure du tube digestif ou métentéron (figure
5.1). Le bord supérieur de ce pli, ou pli amniotique caudal, rejoint ses correspondants crânial
et latéraux.
C) LES PLIS LATERAUX
Ils délimitent latéralement l'embryon de ses annexes en reliant les plis céphalique et
caudal (figures 5.1 et 5.2). Leur bord supérieur, ou plis amniotiques latéraux, rejoignent les
plis céphalique et caudal pour refermer la cavité amniotique "au dessus" de l'embryon. Ce
type d'amniogenèse ou pleuramnios est caractéristique des ruminants, des équins et des
carnivores. Les primates et les rongeurs présentent une amniogenèse différente ou
schizamnios (cf. chapitre VI).
5.2. L'EVOLUTION DES FEUILLETS
La figure 3.16 représente les dérivés des trois feuillets fondamentaux.
5.2.1. L'ectoderme
L'ectoderme est le feuillet le plus externe du disque embryonnaire. Ses deux dérivés
les plus importants par leur développement sont le système nerveux central et l'épiderme
(figure 5.10).
La formation du système nerveux central ou neurulation commence par le
positionnement d'une plaque neurale en avant de la ligne primitive. Cette plaque neurale
apparaît sous l'action inductrice de la chorde dorsale mise en place lors de la gastrulation.
Elle est constituée de cellules ectodermiques qui se différencient progressivement en
neurectoderme. Cette plaque neurale se creuse longitudinalement pour se transformer en
gouttière neurale dont les bords libres portent le nom de crêtes neurales (figures 5.3 et 5.4).
La gouttière neurale se referme pour se transformer en tube neural. Cette fermeture débute en
région médiane et progresse ensuite vers les pôles crânial et caudal (figure 5.5 et 5.6). Le
tube neural reste ouvert à ses deux extrémités par les neuropodes crânial et caudal. Ces
derniers se refermeront plus tard(1).
Lors de la fermeture du tube neural, les cellules des crêtes neurales s'isolent sous la
forme de petits amas à l'origine des ganglions rachidiens (figure 5.4).
(1) La persistance du neuropode antérieur entraîne un encéphalocoele (défaut létal) et celle du neuropode
postérieur une spina bifida (défaut létal ou incapacitant)

Dans le même temps, l'ectoderme participe à l'édification des sillons et des arcs
branchiaux, des bourgeons faciaux (figure 5.5) et des trois placodes (figures 5.7 et 5.8). Ces
différentes structures sont les ébauches d'organes aussi divers que les organes des sens (ouïe,
odorat, vue), les cavités buccale et nasale, la mandibule, l'appareil hyoïdien, les osselets de
l'oreille, l'adénohypophyse ou certaines glandes salivaires (voir le cours d'organogenèse).
Au niveau du tronc, l'ectoderme participe à la formation des bourgeons des membres.
Les bourgeons des membres antérieurs apparaissent avant ceux des membres postérieurs. Ce
décalage chronologique est particulièrement marqué chez les marsupiaux. Ces derniers
naissent à l'état fœtal et doivent rejoindre la poche marsupiale pour y poursuivre leur
développement. A ce stade, leurs membres antérieurs sont parfaitement fonctionnels alors
que les postérieurs sont à peine ébauchés (figure 5.9).
5.2.2. L'endoderme (embryonnaire)
L'endoderme embryonnaire est le feuillet le plus interne du disque embryonnaire
(figure 5.11). Ses principaux dérivés sont :
a) l'épithélium qui recouvre le tube digestif depuis l'œsophage
jusqu'au rectum
b) l'épithélium des voies respiratoires depuis le larynx jusqu'aux
alvéoles
c) les parenchymes hépatique et pancréatiques (endocrine et
exocrine).
L'endoderme embryonnaire est donc à l'origine des cellules fonctionnelles de la
digestion et de la respiration (figure 5.11). Il forme également l'épithélium vésical car la
vessie dérive, comme le rectum, de la partie terminale du tube digestif.
5.2.3. Le mésoderme
Le mésoderme est le feuillet médian de l'embryon trilaminaire. Il est à l'origine de
tout ce qui n'est ni ectodermique, ni endodermique. Ses dérivés constituent l'essentiel de la
masse corporelle et comprennent les systèmes :

a) musculo-squelettique (os, cartilages, muscles striés, lisses et cardiaque)
b) uro-génital (reins, uretères, gonades, voies génitales)
c) circulatoire (circulations sanguine et lymphatique)
Tous les dérivés endodermiques et ectodermiques sont doublés d'un feuillet
mésodermique qui assurera le développement des structures de soutien du futur organe (figure
5.12).
Au niveau du tronc, la lame mésodermique primitive qui sépare l'ectoderme de
l'endoderme primitif (cf. chapitre III) se divise en trois bandes longitudinales de part et d'autre
de la chorde (figure 5.13) :
a) le mésoderme paraxial qui se segmente en somites à l'origine du squelette
axial, de la musculature et du derme de la peau.
b) le mésoderme intermédiaire à l'origine de l'essentiel des systèmes urinaire et
génital.
c) le mésoderme latéral qui se dédouble en deux feuillets.
Le feuillet externe ou mésoderme pariétal (ou somatique) borde l'ectoderme
embryonnaire et extraembryonnaire pour former la somatopleure extra- et intra-embryonnaire
(figure 5.14).
Le feuillet interne ou mésoderme viscéral (ou splanchnique) qui borde l'endoderme
intra- et extra-embryonnaire pour former la splanchnopleure intra- et extra-embryonnaire
(figure 5.14).
Entre ces deux feuillets se trouve une cavité le plus souvent virtuelle : le cœlome qui,
lors de la délimitation de l'embryon, se divisera en cœlome extra- et intra-embryonnaire. Le
cœlome intraembryonnaire est à l'origine des trois grandes cavités internes et de l'endothélium
qui les tapisse :
- la cavité péricardique et le péricarde
- la cavité pleurale et la plèvre
- la cavité péritonéale et le péritoine.
Le mésoderme pariétal fournira également le mésoderme des membres en
collaboration avec le mésoderme paraxial.
Au niveau de la tête, le prolongement céphalique de la chorde (ou lame précordale)
engendre la formation des arcs branchiaux et la mise en place des somites céphaliques
partiellement segmentés. Ces derniers sont à l'origine des éléments mésodermiques de la tête
(squelette, muscles, cartilages, ...).
1
/
4
100%