Chapitre 3 : La mitose Elle correspond à la phase M ou phase de

Chapitre 3 : La mitose
Elle correspond à la phase M ou phase de division cellulaire et inclut les différentes étapes de la
division nucléaire (du noyau de la cellule) ou mitose et de la cytodiérèse ou division du
cytoplasmique. C'est pendant cette phase que le contenu de la cellule dite mère est dupliqué puis
séparé en deux cellules filles génétiquement identiques à la cellule mère. Au niveau moléculaire,
cette phase M est initiée par une cascade de phosphorylations protéiques, provoquée par
l'activation de la protéine kinase MPF ou Mitotic Promoting Factor (voir page précédente : les
cyclines) inductrice de la mitose.Ces phosphorylations protéiques qui se produisent pendant la
mitose sont responsables de la plupart des changements morphologiques qui se produisent
pendant cette phase : condensation des chromosomes, rupture de l'enveloppe nucléaire,
fragmentation du RE (réticulum endoplasmique) et de l'appareil de Golgi, perte de la capacité
d'adhérence des cellules les unes aux autres ainsi qu'à la matrice extracellulaire, modifications du
cytosquelette pour produire des mouvements organisés des chromosomes vers les deux nouvelles
cellules, bref une réorganisation complète de la cellule. Avant le déclenchement proprement dit de
la phase M et la ségrégation (séparation) des chromosomes, on observe leur condensation. Les
schémas suivants des différentes phases représentent un exemple théorique d'une cellule à 3
chromosomes.
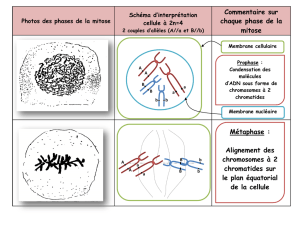
1. La prophase : * pro : du grec pro- , préfixe qui peut signifier : devant, avant, mais aussi
favorable à, pour, à la place de
* phase : du grec phasis [phase, -phasie] : qui ici le sens de "chacun des
aspects successifs d’un phénomène".

* Chromosomes : la chromatine qui était diffuse se condense et forme des chromosomes de plus
en plus individualisés. Ces chromosomes sont à 2 chromatides sœurs, car ils se sont dupliqués
pendant la phase S (voir page précédente - "Le cycle cellulaire"). Les chromatides sont attachées
par une séquence d'ADN particulièrement condensée au niveau du centromère.
* Fuseau : les nombreux microtubules cytoplasmiques qui constituent le cytosquelette dans la
cellule en interphase (c'est-à-dire qui ne se divise pas) se dépolymérisent en molécules de
tubuline, permettant la formation progressive de l'un des éléments fondamentaux de la mitose : le
fuseau mitotique (que l'on appelle aussi le fuseau achromatique). Comme le montre le schéma, ce
fuseau commence à se former à l'extérieur du noyau, en associant les microtubules
cytoplasmiques et des protéines spécifiques.
Ces microtubules sont attachés aux centrosomes par leur extrémité "-" (moins), alors que
l'extrémité "+" (plus) est dynamiquement instable, pouvant passer subitement d'une croissance
uniforme à un raccourcissement rapide par dépolymérisation. Si deux microtubules émanant de
centrioles différents entrent en contact, il est possible qu'ils se stabilisent par des liaisons
transversales. En même temps, les microtubules s'orientent de façon à repousser les pôles du
fuseau. Donc en réalité, le fuseau est formé par 2 moitiés fuseaux.
* Nucléole : Le matériel nucléolaire disparaît progressivement.
* Centrosomes et asters : les centrosomes, dont la duplication s'est faite pendant les stades G1 et
G2, s'éloignent l'un de l'autre et vont devenir de véritables centres mitotiques qui organisent des

microtubules :
certains
formeront une
structure
étoilée, appelée
pour cette
raison l'aster ;
d'autres vont
développer les
fibres du fuseau
entre les
centres
mitotiques : les
microtubules
polaires.
* Kinétochores : en fin de prophase, des structures spécialisées à trois couches appelées
kinétochores, formés de complexes protéiques spécialisés, se développent et s'attachent dans la
région du centromère. Il y a un kinétochore pour chaque chromatide. Ils vont jouer un rôle
primordial au moment de la séparation des chromatides. Les microtubules kinétochoriens, insérés
dans le kinétochore se développent progressivement et, dans la prophase tardive (entre prophase
et prométaphase), ils vont progressivement s'attacher aux microtubules du fuseau ou microtubules
polaires.
2. La prométaphase : * méta : du grec meta exprimant la succession, le changement,
signifiant également plus loin, à côté de, entre, avec.
* Membrane nucléaire : le premier événement majeur de la prométaphase est la désagrégation, en
moins de 30 secondes, de la membrane nucléaire qui se dissocie en de nombreuses vésicules.
Elles resteront visibles autour du fuseau jusqu'à la reconstitution du noyau. En même temps, les
gros organites se dissocient aussi en vésicules, mais ce phénomène est moins visible (et non
représenté), notamment le réticulum endoplasmique et l'appareil de Golgi.
* Chromosomes et microtubules kinétochoriens : les chromosomes sont maintenant dans le
cytoplasme et sont progressivement agités de mouvements désordonnés. Les microtubules
polaires ont pu pénétrer dans l'espace du noyau et se fixent sur les kinétochores modifiés à cet
effet. Sur les deux kinétochores d'un même chromosome doivent se fixer impérativement deux
microtubules provenant chacun d'un pôle différent, sous peine de mauvaise ségrégation (il y aurait
un chromosome de trop dans une cellule fille alors qu'il manquerait dans l'autre).
Les microtubules polaires qui viennent de s'accrocher aux kinétochores sont maintenant appelés
microtubules kinétochoriens. Ce sont eux qui exercent des tensions sur les chromosomes
auxquels ils sont attachés, entraînant les mouvements chromosomiques observables.
3. La métaphase :

A cet instant de la mitose,
le rôle essentiel des
microtubules
kinétochoriens est d'aligner
tous les chromosomes sur
un plan virtuel, situé au
milieu du fuseau et appelé
pour cette raison plan
équatorial. La tension
exercée sur les 2
kinétochores d'un même
chromosome par les
microtubules
kinétochoriens est
constante et équilibrée.
Cette phase est
intéressante à plus d'un
titre car tous les chromosomes sont sur un même plan ; c'est pourquoi elle est utilisée pour réaliser
les caryotypes. A noter qu'à la fin de la métaphase, les chromosomes ont atteint leur maximum de
condensation.
(*) Caryotype : Génétique, gynécologie obstétrique, pédiatrie - N. m. * caryo, karyo : du grec
karuon [cary(o)-, kary(o)-], noix, noyau ; * type : du grec tupos [-type, -typie, -typique], empreinte,
modèle. Un caryotype est la présentation photographique ou dessinée du nombre et de la forme
des chromosomes. Le caryotype est caractéristique de l'espèce. Principales étapes pour la
réalisation d'un caryotype : photographie d'une cellule somatique (pas une cellule reproductrice) en
division (ou en mitose). Cette cellule a été volontairement bloquée en métaphase de mitose, alors
que les chromosomes dupliqués sont pourvus de deux chromatides identiques. Une fois la photo
développée, les chromosomes sont découpés par un personnel qualifié (cytogénéticien, -enne)
puis classés par taille et mis en place par paires homologues. Un exemple de caryotype normal
Tout individu possède normalement 46 chromosomes, dont deux chromosomes sexuels : XX pour
les femmes, XY pour les hommes. Les caryotypes normaux sont donc : 46, XY pour un homme,
46, XX pour une femme. Autre exemple : fillette atteinte de trisomie 21 : 47, XX, 21+ soit un total
de 47 chromosomes, XX pour une fille et 21+ indique que c'est le chromosome 21 qui est
surnuméraire. Caryotype d'un garçon atteint de trisomie 21 Actuellement, les opérations
fastidieuses de découpage ne se font pratiquement plus et le classement est réalisé par un
ordinateur.
Dans le fuseau métaphasique, responsable de la stabilité apparente des chromosomes sur le plan
équatorial, il y a en fait un remaniement incessant des microtubules. Des monomères (molécules
de tubuline) viennent constamment se rajouter à leurs extrémités plus (+), c'est-à-dire à l'équateur
du fuseau, alors que d'autres sont régulièrement perdus aux extrémités moins (-), c'est-à-dire aux
pôles du fuseau.
4. L'anaphase : * ana : du grec ana qui a de nombreux sens : de nouveau, en sens contraire,
en haut, en arrière, à l'écart

L'anaphase ne dure que
quelques minutes, démarre
brusquement et est marquée par
:
* la séparation des deux
chromatides sœurs de chaque
chromosomes,
* leur mouvement vers les pôles
(chaque chromatide sœur vers
un pôle différent), à une vitesse
moyenne de 1 μm par minute
* l'allongement du fuseau et de la
cellule, pour permettre la
séparation future en deux
cellules filles.
Lorsque les nouveaux
chromosomes fils (les
chromatides de départ contenant
chacune une molécule d'ADN
double brin) se déplacent vers
les pôles opposé du fuseau, il est
facile de voir que ce mouvement est dû à la rupture des microtubules au niveau du kinétochore de
chaque chromosome fils. Lorsque les kinétochores se séparent, les bras des chromosomes
avancent péniblement et passivement. Cette rupture est probablement due aux microtubules
kinétochoriens qui se raccourcissent, tandis que les microtubules du kinétochore glissent sur les
microtubules polaires. Ces mouvements caractérisent une première partie de l'anaphase appelée
anaphase A.
Ensuite, pendant l'anaphase B, les pôles du fuseau s'éloignent, par suite de l'action de certains
microtubules polaires qui contribuent ainsi à la séparation des chromosomes fils. Les microtubules
polaires contiennent de la dyénine ou "protéine du mouvement", protéine qui est aussi associée
aux microtubules des cils et des flagelles. Il est donc possible que le mouvement vers les pôles
s'effectue comme le battement des cils et des flagelles des Eucaryotes.
Des analyses ont montré que l'hydrolyse d'une vingtaine de molécules d'ATP seulement suffit pour
permettre aux chromosomes de se déplacer de la plaque équatoriale vers les pôles.
Autre phénomène important et étonnant : le vent polaire ou force d'exclusion astrale.
Des expériences de microsection des bras d'un chromosome ont montré que, si la partie
kinétochorienne attachée au microtubule se dirigeait bien vers le pôle, les bras étaient repoussés
du pôle. Cette force, qui a été appelée force d'exclusion astrale ou vent polaire, pourrait résulter
d'une poussée par les extrémités en croissance des microtubules qui s'assemblent
continuellement au niveau des pôles. On pense maintenant que cette force d'exclusion astrale
pourrait être à l'origine du maintien des chromosomes sur le plan équatorial pendant la métaphase.
5. La télophase : * télo : du grec teleos [téléo-, télo-], fin (comme fini).
 6
7
8
6
7
8
1
/
8
100%











){kind=link}
){kind=link}