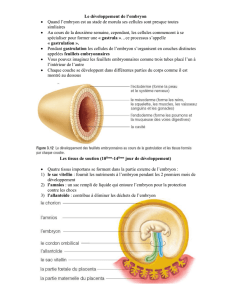
Les mammifères

EMBRYO : mammifères - 2010 Page 1
Les mammifères
Nidation : Le blastocyste va se fixer sur la paroi utérine et il y aura création d’un placenta.
Gastrulation puis chez la souris, il y a une étape particulière de retournement.
Organogenèse (avec neurulation): quasiment tous les organes sont prêts sauf le système reproducteur.
Phase de croissance en taille et poids.
Naissance : fin du développement embryonnaire.
Œuf
Structure d’œuf : ovocyte II, zone pellucide : constituants glycoprotéiques plaqués contre la membrane formant
une coque de protection entourée de cellules folliculaires. L’œuf étant alécithe, il va puiser l'énergie nécessaire à
la segmentation dans les cellules folliculaires puis au cours du temps il y aura perte de ces dernières
placentation.
Après fécondation : achèvement de la méiose, les noyaux deviennent des pronucléi ne fusionnant que tardivement.
Segmentation
(1p6)
Très lente
Se déroule de l’oviducte jusqu’{ l’utérus ;
Divisions asynchrones : 1ère division par clivage méridien, 2ème selon le plan méridien pour un blastomère et
équatorial pour l’autre (segmentation rotationnelle).
Expulsion de l’ovocyte dans les trompes. Rencontre avec un spermatozoïde. Début de la segmentation. Formation
d’une blastula pendant que l’embryon se rapproche de l’utérus, phénomène s’accompagnant d’une perte des
cellules folliculaires mais conservation de la zone pellucide (dont il faudra se séparer pour s’implanter).
Au stade 8 cellules : phénomène de compaction : les blastomères se plaquent les uns contre les autres formant
un amas cellulaire compact sphérique (coopération + stabilité métabolique).
Formation de jonctions serrées entre les cellules pour lutter contre les chocs par exemple sachant que la zone
pellucide est fragile, on abouti donc à une forme avec une plus grande stabilité. Il y a une coopération métabolique
grâce aux jonctions, elles vont donc pouvoir se faire passer des choses de l’une { l’autre (signaux, nutriments,
énergie…), ça sert { rentabiliser au maximum le peu d’énergie que l’on a (absence de vitellus et perte des cellules
folliculaires) (2p6)
Stade 16-32 blastomères = morula (/!\ce n’est pas une étape particulière) ; puis blastocyste primaire (avec
blastocœle) constitué de trophectoderme (périphérique qui donnera le placenta) + bouton embryonnaire
(organo-formateur ; en" battant de cloche" chez la souris, en "massif" plaqué contre le trophoblaste chez l’Homme)
Le blastocyste primaire va perdre toutes les cellules folliculaires et se dégager de la zone pellucide éclosion et
implantation dans l’utérus (polarité définie). Pour casser la coque protéique (zone pellucide), il va falloir que l’œuf
sécrète des enzymes.
Implantation utérine
Implantation utérine : fixation du blastocyste par des cellules du trophectoderme en contact avec le tissu
maternel. (3p6)
Le placenta est une annexe embryonnaire formée pour moitié de trophectoderme (c'est-à-dire de cellules
embryonnaires) et l'autre moitié de cellules maternelles. C'est une zone d'échange par le sang entre l'embryon et
la mère hémotrophie (Il faut donc que les deux parties du placenta soit très vascularisées)
Au moment de l'implantation, dégradation partielle de la paroi utérine. Les éléments qui en sont issus servent
d'énergie pour l'embryon Histotrophie (phase très courte).
Ce sont les cellules du trophectoderme du côté du bouton embryonnaire (polarité définie) qui réagissent avec la
paroi utérine.

EMBRYO : mammifères - 2010 Page 2
/!\Les mammifères sont les seuls animaux à faire une différenciation cellulaire avant gastrulation : cellules
spécialisées dans la blastula pour former l'embryon (bouton embryonnaire). Il y a déjà deux populations
cellulaires distinctes (différent de chez l'oursin).
Mise en place de l’endoblaste = hypoblaste : couche de cellules aplaties sur face inférieure (ventrale) du bouton
embryonnaire (hors de l’embryon) qui va proliférer et se refermer formation d'un sac vitellin vide.
Origine :
- délamination (différent de gastrulation) du bouton embryonnaire chez certains mammifères dont l’homme
- prolifération de cellules trophoblastiques chez d’autres dont le rat.
L’hypoblaste prolifère pour doubler l'intérieur du blastocœle appelé alors lécithocœle (= cavité homologue d’un
sac vitellin mais sans vitellus) tandis que le mésoblaste extra-embryonnaire se met en place.
Les annexes extra-embryonnaires
Formées à partir de cellules issues du blastocyste mais ne servant pas à la formation de l'embryon et est cependant
indispensable à sa survie. (1p9)
1. Placenta
Rôles hormonal, d’ancrage de l’embryon dans les voies génitales maternelles, et permet les échanges
mèreembryon (O2, eau, sel, protides, glucides, lipides, vitamines, hormones, anticorps, des médicaments et des
virus) et embryonmère (CO2, eau, urée, anticorps, des déchets et des hormones).
Capable d’agir comme une vraie glande en produisant des hormones pour le bon développement et pour la
signalisation de la présence de l’embryon3 sources d’hormones : la mère, le placenta et l’embryon.
Partie maternelle = muqueuse utérine hypertrophiée et très vascularisée + partie embryonnaire = chorion =
trophoblaste doublé par la somatopleure extra embryonnaire, le chorion développe des villosités (qui seront
vascularisées) à sa surface et est doublé par des parois d’annexes. Chez la souris et l’homme, le chorion assure
toutes les fonctions placentaires
Les différents types de placenta (2-3p9)
Selon les implications des annexes, placentation
- Vitello-chorionique (ou allanto-vitello-chorionique) contact chorion-vésicule vitelline (+/- allantoïde)
- Allanto-chorionique = vascularisation chorionique partiellement d’origine allantoïdienne + contact
éventuel entre chorion et l’allantoïde
Selon si l’utérus et le système vasculaire maternel se conservent ou non intacts
- Indécidués : pas de muqueuse utérine éliminée, ne créant pas d’hémorragie { la naissance
- Placenta indécidué épithélio-chorial, diffus : simple accolement entre les structures maternelles et
embryonnaire.
- Placenta indécidué conjonctivo-chorial (ou syndesmochorial), cotylédonaire (plein de petits
boutons tout autour de l’embryon) = corrosion d’épithélium utérin par endroits (donc échanges
mère-embryon plus intimes car moins de barrière de cellules à traverser)
- Décidués : muqueuse utérine éliminée, hémorragie à la naissance (cas de l’Homme)
- Placenta décidué endothélio-chorial, zonaire : lésion du conjonctif utérin mais pas des vaisseaux
maternels (donc contact direct tissus embryonnaires/vaisseaux maternels)
- Placenta décidué hémo-chorial, discoïdal : utérus corrodé avec attaque des parois des vaisseaux
maternels et formation de lacunes sanguines (donc échanges encore + favorisés, mais pas de mélange
des sangs mère/embryon)
2. Amnios
Cavité emplie de liquide (riche en fructose et hormones) continuellement absorbé par l’embryon par voie buccale
et résorbé vers l’allantoïde par les reins. Etabli par plissement, cavitation (Homme) ou processus mixte (souris).
Non vascularisé, peut (selon le mammifère) être en contact avec le chorion, ne participe pas à la formation du
placenta. Rq: tous les vertébrés la possède.

EMBRYO : mammifères - 2010 Page 3
3. Vésicule vitelline
Rôle par sa vascularisation importante (et peu ou pas de fonction nutritive). Régressée chez les mammifères (de
plus en plus des marsupiaux aux primates), peut participer à la formation (en renforçant le chorion
(structurellement parlant en apportant une autre couche de cellule en s’accolant { lui) vascularisation(en
détournant des vaisseaux sanguins) du placenta ou; Chez souris et homme : +/- { l’origine de cellules sanguines, de
plus, chez l’homme : rôle respiratoire au début des stades d’organogenèse.
4. Allantoïde
Peut être { l’origine de tout ou une partie de la vascularisation du chorion. L’importance et la participation { la
formation du placenta varie selon le mammifère ;
/!\chez la souris et l’homme : allantoïde vestigiale = atrophié et vide surtout vaisseaux irriguant chorion.
Amniogenèse
Avant ou en même temps que la gastrulation, plusieurs modalités de formation selon animaux (1p7) :
1) par plissement : bouton embryonnaire "découvert", s’enfonce sous les replis (lapins, les carnivores, les
ongulés et les primates primitifs).
Déformation de la blastula avec 2 replis côté antérieur et postérieur se soudant entre eux. La cavité ainsi fermée
au-dessus du bouton embryonnaire est l'amnios. Mise en place de l'hypoblaste sac vitellin. Mise en place du
mésenchyme extra-embryonnaire.
2) par cavitation : entre la zone la plus externe cytotrophoblastique et ectoblaste du bouton embryonnaire
formation de vacuoles confluent et donnent une cavité amniotique (insectivores, chiroptères=chauve-souris et
primates dont l’homme).
3) par cyste ectochorial : processus mixte (rongeurs dont souris).
Du mésoblaste forme cœlome extra-embryonnaire, lécithocœle représentera la vésicule vitelline, vésicule
allantoïdienne est ébauchée.
Fin de l’amniogenèse = début de la gastrulation.
Le cœlome coupe en 2 la vésicule vitelline d'où l'obtention de lécithocœle primaire et secondaire.
Gastrulation des oiseaux (identique chez l’Homme)
Début = épaississement d’une zone dû { la migration (convergence) de cellules mésodermiques et
endodermiques. Toute la gastrulation se fait à partir de la ligne primitive. C'est là où affluent les cellules qui
descendent dans le blastocœle et qui ne se rassemblent en feuillet que dans une zone Nœud de Hensen.
Allongement du blastoderme (=environnement extérieur de l’embryon) = Immigration en profondeur de
l’endoderme suivi du mésoderme dans le blastocœle selon la ligne primitive (marque l'axe antérieur-postérieur
de l'embryon) terminée { l’extrémité antérieure par le nœud de Hensen. C'est l'hypoblaste (= structure épithéliale
ventrale, future paroi du lécithocœle) de la blastula qui induit la formation de la ligne primitive. Et divergence
d’endoderme et des mésodermes extra-embryonnaire et embryonnaire.

EMBRYO : mammifères - 2010 Page 4
2 phases dans la gastrulation :
1) Insertion de l’endoderme dans l’hypoblaste forme ébauche du TD et repousse hypoblaste en extra-
embryonnaire ; puis la migration du mésoblaste dans le blastocœle commence dans la moitié postérieure et migre
vers la partie antérieure. La migration antérieure de toutes ces cellules a soulevé la région antérieure de l'épiblaste
et résulte en la formation du bourgeon de la tête.
2) La ligne primitive devient maximale puis recule/régresse vers l’arrière, avec mise en place de la corde par
l’enfoncement de l’endoderme et des somites. Formation du prolongement céphalique (invagination du
mésoderme formant la corde au niveau de la tête). Ce qui reste de l'épiblaste après ces migrations cellulaires, les
cellules de surface, constitue l'ectoderme. (2-3p7)
Conséquence d’une gastrulation en 2 étapes : décalage avec les parties les plus postérieures. Début d’organogenèse
dans la partie antérieure (avec installation du système nerveux au dessus de la corde induction par les cellules
de la corde pour internaliser l’ectoderme) alors que la partie postérieure en est encore à la gastrulation.
/!\La gastrulation est considérée comme achevée avant la disparition totale de ligne primitive.
DIFFERENCES CHEZ L’HOMME :
- A partir de la zone postérieure, il y a formation du nœud postérieur. Plus prés du centre formation du
nœud de Hensen, et entre les deux nœuds : formation de la ligne primitive.
- La corde s’ouvre ventralement et se soude transitoirement { l’hypoblaste puis s’isole en cordon plein
quand les cellules endodermiques s’insèrent dans l’hypoblaste. (2p8)
Mouvements morphogénétiques analogues { ceux de l’oiseau ; avec endoderme et continuité avec l’hypoblaste du
lécithocœle, corde en prolongement céphalique tubulaire.
A la fin de la gastrulation, les somites (= blocs segmentés installés par pairs de part et d’autres de la corde qui
donnent les muscles, le cartilage et le derme) commencent à se métamériser, en même temps que se forme
l’ébauche cardiaque.
Organogenèse
Neurulation : En fin de gastrulation : l’aire ectoblastique au-dessus de la corde est induite à former la
plaque neurale, qui va s’épaissir, se plier en gouttière neurale, puis se fermer en tube neural, vers le
milieu puis antérieur et postérieur, tandis que régresse la ligne primitive
Le mésoderme continue à se découper en
- somites
- pièces intermédiaires
- lames latérales creusées en cœlome
Les cellules germinales primordiales migrent.
TD se referme, et l’embryon s’isole des annexes par un jeu de replis : il n’est plus relié que par le cordon
ombilical contenant les pédicules vitellin et allantoïdien.
Formation de :
- modelage corporel
- système circulatoire
- bourgeons des membres antérieurs
puis postérieurs
- foie, glandes annexes digestives
- ébauches sensorielles
- cerveau, ganglions
- poumons
- dents
- gonades, mésonéphros (reins primitifs).
Chez la souris : inversion de courbure d’embryon puis organogenèse suivie d’une semaine surtout de
croissance
Chez l’homme : morphologie quasi-définitive { 2 mois = fin de l’embryogenèse, (+ placenta vers 1 mois),
puis phase de croissance et maturation fonctionnelle progressive des organes = développement fœtal ;
naissance vers 266jours) puis formation de l’adulte après la naissance.
1
/
4
100%