Corrigé exercices PROCREATION (2) Exercice 2 : Les canaux

1
Corrigé exercices PROCREATION (2)
Exercice 2 :
Les canaux déférents conduisent le sperme depuis l’épididyme jusqu’au canal éjaculateur. Leur ligature
empêche l’excrétion des spermatozoïdes. C’est une technique utilisée pour la stérilisation des individus
femelles. L’irradiation des testicules à l’aide de rayons X empêche la spermatogenèse.
La castration ou ablation des testicules prive l’individu de toute possibilité de se reproduire mais aussi prive
l’individu d’hormones mâles. Le testicule (comme l’ovaire) est en effet une glande mixte endocrine
(hormones mâles) et exocrine (spermatozoïdes). La carence en hormones mâles est plus spectaculaire chez
l’animal jeune car elle interrompt la croissance des organes génitaux et empêche l’apparition des caractères
secondaires.
L’injection d’extraits de cellules interstitielles à un animal castré lui apporte les hormones mâles nécessaires
à la croissance de l’appareil génital et à l’apparition des caractères sexuels secondaires. Les hormones
mâles, dont la principale est la testostérone, sont synthétisées par les cellules de Leydig des testicules.
On peut obtenir le même effet endocrinien en greffant un testicule à l’animal.
Exercice 3 :
a) Sur la coupe microscopique, il faut tout d’abord repérer : tissu interstitiel, paroi du tube séminifère et
lumière de ce tube.
Les cellules de Leydig entourent un capillaire sanguin, ce sont les cellules endocrines sécrétrices de
testostérone.
Les cellules germinales occupent la paroi du tube séminifère et montrent pour certaines des figures de
division ; elles sont à l’origine de la formation des gamètes mâles. La spermatogenèse se déroule de façon
centripète dans cette paroi, les spermatogonies diploïdes étant localisées à la périphérie, les spermatozoïdes
étant libérées dans la lumière du tube.
Les cellules de Sertoli sont les grandes cellules associées aux « bouquets » de spermatozoïdes ; ce sont des
cellules qui ont, entre autre, un rôle nourricier pour les cellules germinales.
b) Les animaux étant impubères, les cellules testiculaires sont normalement inactives. On constate que la
LH stimule les cellules endocrines (la production de testostérone) alors que FSH stimule la lignée germinale
et les cellules de Sertoli (spermatogenèse).
c) La colonne de gauche présente les résultats de référence (témoin). Les cellules de rein, de rate, … ne
modifient pas la sécrétion de gonadostimulines : elles ne libèrent pas de signaux chimiques susceptibles
d’interagir avec les cellules antéhypophysaires responsables de la sécrétion de FSH ou de LH. En revanche,
la présence de cellule de Leydig diminue la sécrétion de LH (mais pas celle de FSH) : c’est la mise en
évidence de la rétroaction négative exercée par la testostérone sur son système de commande.
Exercice 4 :
2.
1) GnRH (Gonadotrophin Releasing Hormon) : neurohormone produite par certains neurones
hypothalamiques de façon pulsatile et stimulant l’hypophyse qui libère FSH et LH.
2) LH (hormone lutéinisante) : libérée par l’antéhypophyse de manière pulsatile, le pic de LH provoque
l’ovulation puis après celle-ci, la LH déclenche la transformation du follicule de De Graaf en corps jaune

2
(chez l’homme la LH stimule le développement des cellules interstitielles de Leydig et la production de
testostérone)
3) FSH (hormone folliculostimulante) : produite par l’adénohypophyse. La FSH provoque la croissance des
follicules de De Graaf notamment en favorisant la multiplication des cellules de la granulosa et des thèques
à l’origine de la production d’oestrogènes. (Chez l’homme, la FSH stimule la spermatogenèse par
l’intermédiaire des cellules de Sertoli).
4) Oestrogènes : sont produits par la thèque interne et la granulosa du follicule ovarien. Les oestrogènes
agissent sur la prolifération des cellules de la muqueuse utérine, sur la sécrétion des glandes du col utérin.
5) Progestérone : hormone produite par le corps jaune favorisant la formation de la dentelle utérine et la
sécrétion de ses glandes ainsi que la sécrétion des glandes du col utérin, elle provoque le développement des
glandes mammaires, une augmentation de la température corporelle et le silence utérin (pas de contraction
du myomètre).
3. La substance 4 (les oestrogènes) exerce d’abord une rétroaction négative puis une rétroaction positive
(selon sa concentration).
Exercice 5 :
Partie I : Régulation du cycle sexuel féminin
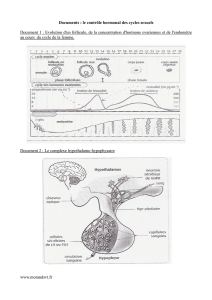
Question 1 :
L’expérience 1 du Document 1 nous indique que l’antéhypophyse stimule l’ovaire et le développement
utérin. Elle participe également à la réalisation du cycle sexuel.
L’expérience 2 permet d’affiner les conclusions de l’expérience n°1. En effet, elle nous montre que l’action
hypophysaire se fait par voie hormonale et qu’elle ne se fait pas directement sur l’utérus. C’est donc l’ovaire
qui, stimulé par l’hypophyse, permet le développement de l’utérus et ce par voie hormonale.
L’expérience 3 nous apprend qu’une hypophyse intacte mais privée de communication avec l’hypothalamus
n’assure plus son rôle. Cette observation, associée à celle de la relation anatomique entre hypothalamus et
hypophyse, nous permet de conclure que l’activité de l’antéhypophyse est due aux sécrétions neuro-
hormonales de l’hypothalamus. De plus, ce dernier est soumis au contrôle des centres nerveux supérieurs
qui peuvent, comme le montre l’observation n°4, l’inhiber ce qui retentit sur l’ensemble du fonctionnement
cyclique de la physiologie sexuelle féminine.
Le Document 2 nous suggère que les ovaires, par leurs sécrétions hormonales, freinent généralement
l’activité hypophysaire. En effet, l’ovariectomie provoque une hypertrophie de l’hypophyse, signe d’un
fonctionnement anormalement élevé. De manière très simpliste, cette action dépend de la présence ainsi que
de la concentration des hormones gonadiques, comme le montrent les données du tableau.
- La période 1 traduit l’hypersécrétion hypophysaire en absence d’ovaires
- Des trois autres périodes, il se dégage que les œstrogènes seuls et à faible concentration
freinent l’antéhypophyse (période 2) mais stimulent cette dernière à forte dose (300 pg/mL, période
3). Cependant la présence des œstrogènes à forte concentration et de la progestérone inhibe les
sécrétions antéhypophysaires qui concernent l’ovaire (période 4).

3
Question 2 :
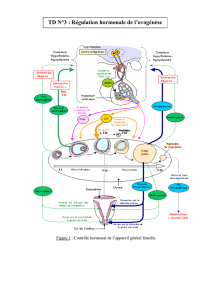
La régulation du cycle sexuel féminin met en jeu plusieurs structures (ovaires, hypophyse, hypothalalamus,
et dans une certaine mesure, des centres nerveux supérieurs) ; à l’exception de cette dernière éventualité,
tous communiquent par des hormones.
- l’hypothalamus sécrète une neurohormone qui stimule l’hypophyse
- cette dernière, par la production de FSH et de LH (gonadostimulines) active les ovaires
- les hormones ovariennes exercent pour leur part une rétroaction négative ou positive sur l’hypophyse
et stimulent le développement de l’utérus
- la boucle de régulation entre système hypothalamo-hypophysaire permet la mise en place d’un cycle
sexuel chez la femme.
Partie II : La contraception
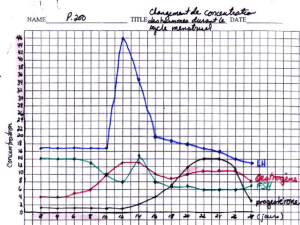
Les graphiques du Document n°3 montrent que la prise d’un pilule normo-dosée annule les sécrétions
cycliques des hormones ovariennes observées antérieurement de 0 à 28 jours, ainsi que l’apparition des pics
de FSH et de LH qui marque le milieu du cycle.
Le Document n°4 permet de confirmer ce qui a été dit dans la partie I à savoir que l’hypothalamus stimule
l’hypophyse. Ce Document nous apporte deux informations supplémentaires : la première est que la neuro-
hormone se nomme GnRH, la seconde que les sécrétions hypophysaires et hypothalamiques sont pulsatiles.
Les données du Document n°5 sont à mettre en relation avec celles du tableau de la partie I. Elles nous
indiquent clairement que les oestrogènes (ici l’oestradiol) freinent la sécrétion de LH (rétroaction négative)
mais qu’au-delà d’une certaine concentration, déclenchent une sécrétion brutale et abondante de LH. Cette
expérience reconstitue la phase folliculaire du cycle où l’on observe une lente augmentation du taux
plasmatique d’oestrogènes jusqu’à ce qu’une accumulation se traduise par un taux plasmatique élevé ayant

4
pour conséquence un effet positif sur la sécrétion de LH ; celui-ci prend la forme d’un pic qui à son tour
déclenche l’ovulation.
La prise d’une pilule normo-dosée a pour but d’éviter l’ovulation. Il faut pour cela annuler le pic ovulatoire
de LH et donc l’apparition d’une forte concentration d’oestrogènes qui est la conséquence de la maturation
d’un follicule IIIaire et son évolution en follicule mûr. Cette maturation est la conséquence de la stimulation
de l’ovaire par la FSH. En diminuant son taux (par l’apport d’une faible dose d’oestrogènes que l’on associe
à de la progestérone), on évite le développement folliculaire et donc l’ovulation.
L’effet du contraceptif est de renforcer le rétrocontrôle négatif qu’exercent normalement les hormones
ovariennes sur le complexe hypothalamo-hypophysaire.
Exercice 6 :
La grossesse représente le développement de l’embryon puis du fœtus dans l’utérus maternel, qui lui doit
aussi se développer. Comment peut-on expliquer qu’une molécule contragestive, le RU486, puisse stopper
une grossesse débutante ?
- Importance de la progestérone pendant la grossesse
Document 1 : Les expériences du Document 4 ont été résumées dans un tableau. Celles-ci ont été réalisées
sur des lapines impubères c’est-à-dire des lapines qui ne sécrètent plus d’hormones ovariennes naturelles ( à
savoir les œstrogènes et la progestérone).
Si on compare les lots 1 et 2, on constate que l’œstradiol présent en 2 conduit à une faible augmentation de
la paroi utérine mais la dentelle utérine ne se forme pas.
Si on compare les lots 3 et 2, on constate que la présence simultanée d’œstrogènes et de progestérone
entraîne un développement important de la paroi utérine et la formation de la dentelle utérine. Cette dernière
permettra la nidation (fixation) et la nutrition de l’embryon.
Document 2 : Le document est un graphique avec en ordonnée le taux de phénolstéroïdes, produit de
l’élimination des œstrogènes le taux de prégnandiol, produit de l’élimination de la progestérone et en
abscisse le temps en jours. Ce document nous montre que le dosage des hormones ovariennes s’effectue
pendant environ 73 jours. On observe un cycle ovarien normal du 1er au 30/09 environ. Le début d’un
nouveau cycle, au 1er jour de l’apparition des règles, est précédé d’une chute importante du taux des
hormones ovariennes.
Les œstrogènes et la progestérone agissent au niveau de l’utérus (voir Document 4) et sont responsables de
la formation de la dentelle utérine.
Dans la 2nde partie du graphique traduisant le second cycle ovarien, on observe un taux important
d’hormones ovariennes. Ceci est le résultat d’une fécondation. Le taux de progestérone passe de 1mg/L à
7 mg/L. Cette hormone a un rôle essentiel dans les transformations de l’utérus nécessaire à une bonne
grossesse.
Document 3 : Le Document 3 nous présente un tableau montrant les résultats correspondant au comptage
des grains d’Ag (révélant la radioactivité) réalisés sur différentes coupes de la muqueuse utérine et du
muscle utérin. L’injection de RU486 marqué lors de la 1ière expérience et l’injection de progestérone
marquée lors de la 2nde expérience montrent la présence de radioactivité dans les cellules de la muqueuse
utérine et dans les cellules du muscle utérin. Ceci montre que la RU486 et la progestérone se fixent sur
l’endomètre et le muscle utérin car ces cellules deviennent radioactives. Ces cellules utérines correspondent
aux cellules cibles de la progestérone.

5
Comment le RU486 entraîne l’apparition des règles et l’expulsion de l’embryon implanté dans l’endomètre
utérin ?
- Mode d’action du RU486
Document 4 : La comparaison des lots 4 et 5 avec le lot 3 montre que des doses croissantes de RU486
entraînent une régression de la paroi utérine puis une disparition de la dentelle utérine. Le RU486 réduit et
finalement annule l’action des hormones ovariennes et plus précisément agit à l’encontre de la progestérone.
Document 3 : La comparaison des lignes 2 et 3 des résultats du tableau montre que l’injection de RU486
puis de progestérone marquée entraîne une fixation de progestérone radioactive 3,5 fois moins importante
sur les 2 types de cellules utérines. Ceci montre que les récepteurs de la progestérone sont bien ceux sur
lesquels se fixe la RU486. Ce dernier se fixe sur les récepteurs de la progestérone et empêche celle-ci de se
fixer. Ainsi ceci explique les résultats des lots 4 et 5 du Document 4. Le RU486 entraîne un moindre
développement de l’utérus.
Document 1 : Il nous présente les formules chimiques de la progestérone et du RU486. Ces 2 molécules
présentent les mêmes structures polycycliques à savoir celui du cholestrol. On peut émettre l’hypothèse que
ces 2 molécules se fixent sur les mêmes récepteurs des cellules utérines. Du fait de cette ressemblance
structurale, le RU486 entre en compétition avec la progestérone au niveau des sites récepteurs de celle-ci.
Cette compétition engendre une moins bonne fixation de la progestérone (Document 3) et le moindre
développement de l’endomètre quand le RU486 est en concentration suffisante (Document 4).
Conclusion : le RU486 entrant en compétition avec la progestérone entraîne une réduction puis supprime
les effets de cette hormone ovarienne. On peut dire que le RU486 est une molécule anti-progestérone.
Utilisé en début de grossesse, le RU486 entraîne la disparition de la dentelle utérine, l’apparition des règles
et l’expulsion de l’embryon en début de gestation. Ainsi est résumé l’effet contragestif du RU486.
Exercice 7 :
1. X : Follicule cavitaire ou tertiaire
Y : Follicule mûr (de De Graaf)
Z : Corps jaune
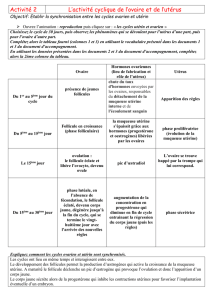
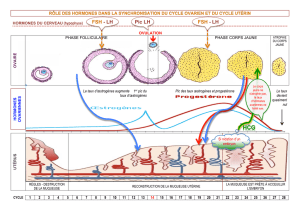
2. Le Document 2a correspond à un cycle ovarien au cours duquel on analyse l’évolution des hormones
ovariennes, œstrogènes et progestérone.
- Au cours de la phase folliculaire, qui début au 1ier jour des règles, il y a maturation d’un follicule
cavitaire (X, Document 1). L’hormone sécrétée est l’œstradiol, elle provient des cellules de la thèque
interne et de la granulosa du follicule. La sécrétion est faible jusqu’au 9ième jour, elle augmente avec
un maximum au 12ième jour (250 pg/mL).
- Vers le 14ième jour, le follicule, devenu follicule mûr (Y, Document 1) expulse son ovocyte par
déchirure de la paroi de l’antrum et rupture du cumulus oophurus.
- A partir des vestiges du follicule se développe un corps jaune (Z, Document 1) dont les cellules
lutéales sécrètent la progestérone alors que la thèque interne continue, à un taux modéré, de sécréter
des œstrogènes. Cette phase dure également 14 jours. Le taux de progestérone atteint un maximum
vers le 21ème jour (16 ng/mL). Si l’ovocyte expulsé n’est pas fécondé, le corps jaune s’atrophie et le
taux de progestérone et d’œstrogènes ne deviennent pas maximaux.
L’épithélium utérin, cible des hormones ovariennes, s’est épaissi pendant la phase folliculaire, a
continué de s’épaissir en se différenciant (dentellisation, vascularisation) au cours de la phase
lutéale. Les très faibles taux d’œstrogènes et de progestérone, conséquence de la dégénérescence du
corps jaune, entraîne son effondrement : ce sont les règles qui marquent le début d’un nouveau cycle.
 6
7
8
9
6
7
8
9
1
/
9
100%