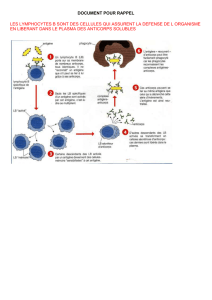
thymus et moelle rouge des os longs.

1
IMMUNITE

2
Sommaire
• Introduction au système immunitaire
• Mécanismes de l’immunité innée et acquise
• Le syndrome d’immunodéficience humaine acquise
• La vaccination

3
Introduction au système immunitaire

4
Introduction
•
I- Introduction
L'immunité désigne l'ensemble des mécanismes développés par un organisme pour se maintenir
en bonne santé.
L'immunité a donc un double aspect:
* l'intégrité de l'organisme (sa cohérence, son fonctionnement intégré; c'est l'immunité au sens
plus ancien mais c'est aussi une ouverture sur un sens très actuel de système de communication
tourné vers l'intérieur, comme nous le verrons)
* et sa défense vis-à-vis des éléments étrangers ou antigènes- éléments susceptibles de
déclencher une réponse immunitaire, après reconnaissance par des cellules immunitaires -
(immunité au sens plus courant).

5
Organisation du système immunitaire
II- Structure du système immunitaire
Le système immunitaire comprend trois types d'organes:
- les organes centraux ou primaires: thymus et moelle rouge des os longs.
-les organes périphériques ou secondaires :ganglions lymphatiques et rate
-les appareils circulatoires (sanguin et lymphatique).
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
1
/
55
100%