CHLOROPLASTES
DEFINITION
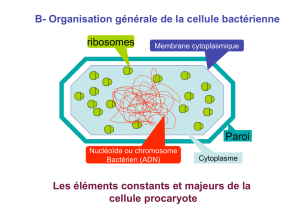
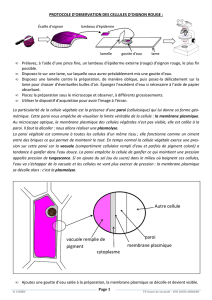
Les chloroplastes sont des organites présents dans le cytoplasme des cellules végétales
eucaryotes. L'ensemble des chloroplastes d'une cellule constitue le plastidome.
Les chloroplastes sont caractérisés par leurs pigments, chlorophylles et caroténoïdes, qui
assurent l'absorption de l'énergie solaire qu'ils transforment en énergie chimique au cours de la
photosynthèse.
1-STRUCTURE ET ULTRASTRUCTURE DES CHLOROPLASTES (voir TP)
1- STRUCTURE : (M.P).
Chez les végétaux supérieurs (Plantae) ils ont une forme ovoïde, et de couleur verte due aux
pigments chlorophylliens.
2- ULTRASTRUCTURE : MET (polycopié p. 133)
2.1 - Observation au MET faible grossissement
L'observation au MET montre que le chloroplaste se présente comme un disque ovoïde ou
lenticulaire de 3 à 10.µm de long et 1 à 2µm de diamètre.
Le chloroplaste est constitué de trois (03) compartiments qui coopèrent étroitement pour
réaliser la photosynthèse:
-L'enveloppe: formée d'une double membrane, une membrane externe et une membrane
interne, délimitant le chloroplaste.
-Les thylakoïdes: réseau membranaire présentant une structure extrêmement ordonnée sous
forme de citernes aplaties plus ou moins longues constituant les thylakoïdes. Les citernes les
plus courtes sont empilées les unes sur les autres comme des pièces de monnaies pour former
les grana (granum au sing.).
-Le stroma : milieu dans lequel baignent les thylakoïdes, riche en protéines solubles. Il
présente un aspect granuleux au MET dû essentiellement à sa richesse en ribosomes.
2.2 - Observation aux MET et MB fort grossissement
a-Au MET :
Les membranes externe et interne de l'enveloppe (6 nm d'épaisseur) et les membranes des
thylakoïdes (6à8 nm d'épaisseur) présentent une structure trilamellaire.
b- Au MEB:
Présence de particules globulaires dans les 3 membranes asymétrie plus importante au niveau
des membranes thylakoïdales.
L'application de la technique de coloration négative a permis de mettre en évidence dans les
membranes des thylakoïdes la présence d'ATP synthétases (appelés anciennement ATPosomes
ou CF0-CF1) dont les sphères, parties hydrophiles de 9nm de diamètre sont orientées vers le
stroma (l'observation est faite au MET fort grossissement).
II-COMPOSITION CHIMIQUE:
1-Isolement de fractions et sous-fractions:
I-1 Fraction
Broyage de feuilles homogénat 2ème culot : chloroplastes
Filtration 2UCD

1-2- Sous fraction
Choc osmotique
+UCD 2UCD
Chloroplaste membrane externe +contenu intermembranaire
Ultrasons
+2 UCD
UCD gradient
saccharose
Dans les couches du gradient de saccharose, on peut reconnaître les thyIakodee à leur couleur verte (riche en
chlorophylles).
2- Résultats de l'analyse chimique:
2-1- Membranes de l'enveloppe et des thylakoïdes (tableau ci-après).
Mbs
Constituants
ENVELOPPE
(Mb externe perméable)
(Mb interne sélective)
(Composition globale)
THYLAKOIDES
(Mb sélective)
LIPIDES
60%
Riche en glycolipides
Pauvre en phospholipides
Riche en sulfolipides
38%
glycolipides
pauvre en phospholipides
riche ensulfolipides
(riche en AG insaturés, fluidité
membranaire plus importante)
PROTEINES
40%
Taux plus faible dans mbrane externe
glycosyltransférases
transporteurs= perméases passives et
actives (contrôlent
le passage des molécules entre stroma et
hyaloplasme)
50%
4complexes
-PSII et PSI (protéines +pigments)
capteur de photons
-chaîne photosynthétique transporteurs
d'e et/ou de H+:
cytochrome ou complexe b6-f
plastocyanine(PC) mobile
plastoqninone(PQ) mobile
-ATPsynthétase
PIGMENTS
Caroténoïdes (faible %)
absence de chlorophylles
12%
-chlorophylles (10%)
-caroténoïdes (211%)
Associés aux PSII et PSI
Contenu intermembranaire
Contenu du stroma
Membrane interne
+ thylakoides
Mb interne
Thyakoïdes
Membrane externe

Fig. 1 Chaîne de transfert des électrons et/ou de protons
Organisation de la membrane tyhlakoidienne des chloroplastes des végétaux supérieurs
Les photosystèmes I et II, ainsi qu’un gros complexe d’oxydoréduction membranaire(dit
complexe b6-f), participe à la capture de la lumière et à la création du gradient de protons qui
est exploité par l’ATP synthétase pour fabriquer l’ATP. La plastocyanine (PC) et la ferrédoxine
(Fd) sont des protéines mobiles transportant des électrons ; la plastoquinone (Q) est un
transporteur d’hydrogène.
Fig 2 : Cycle de Calvin- Bassham Bensson

2-2- Architecture moléculaire des thylakoides (Fig. 1 et polycopié p. 133)
A enlever sur polycopié p.133 la NADP réductase qui est considérée actuellement comme
faisant partie du PSI.
2-3- Contenu de l'espace intermembranaire: Contenu aqueux formé par les constituants qui
transitent entre le stroma et le cytoplasme: CO2, O2, ATP, ADP, H2O, ions, protéines etc.
2-4- Contenu du stroma: Solution aqueuse renfermant: enzymes du cycle de Calvin, acides
aminés, acides gras, oses, nucléotides, ADN, ARN, grains d'amidon, plastoribosomes,
plastoglobules (granules lipidiques), ions Mg++.
III-FONCTIONS (Fig. 1 et polycopié p.13 5)
1-PROTOSYNTHESE.
En présence d'eau et dioxyde de carbone, les végétaux photosynthétiques convertissent
l'énergie lumineuse en énergie chimique et produisent des sucres (amidon). Ce processus appelé
photosynthèse s'accompagne d'une libération d'oxygène.
Lumière
6CO2 + 12H2O C6H12O2 + 6O2 +6H2O + énergie
Elle comporte 02 phases:
1-1- Phase claire ou primaire (fig.1)
- absorption de la lumière par les pigments au niveau des PSI et PSII,
- formation de NADPH+H+
* dégagement d'oxygène
- production d’ATP
Etapes:
-L'énergie lumineuse est captée au niveau de la membrane des thylakoïdes par les
photosystèmes IIet I (PSII et PSI)
-Les chlorophylles des centres réactionnels (chla 680 du PSII et Chla 700 du PSI) s'oxydent
par perte d'e.
-La photolyse de l'eau libère de l’O2 et 2e- qui permettent au PSII de revenir à son état initial.
-Les e- et les H+ sont ensuite transportés par une succession de réactions d'oxydoréduction
grâce à des transporteurs de la chaîne photosynthétique. Le PSI réduit le NADP+ en NADPH2,
grâce à la NADP-réductase, qui en fait partie. Les protons s'accumulent dans l'espace
intrathylakoidal ou lumen, créant un gradient électrochimique. Le retour des protons vers le
stroma se fait par l'ATPsynthétase entraînant la synthèse d'ATP: c'est la
photophosphorylation
1-2- Phase sombre ou secondaire (fig.2)
- Fixation de CO2 grâce à la Rubisco (enzyme Ribulose 1-5 bi phosphate
carboxylaseoxygénase).
- L'ATP et le NADPH2 formés au cours de la phase claire sont utilisés dans le cycle de Calvin-
Bassham- Bensson, pour former des molécules organiques (glucides à 3C), qui seront soit
transformées en glucose puis en amidon qui s'accumule, ou exportés vers le cytosol,
directement après leur formation ou indirectement après dégradation de l'amidon. Dans le
cytosol ils serviront alors de substrat pour les chaînes de biosynthèses d'autres sucres, de
lipides, acides aminés, nucléotides etc. Ils peuvent aussi être oxydé dans les mitochondries et
libérer leur énergie sous forme d'ATP.

2- SYNTHESE DES PROTEINES (polycopié p.. 137) comparaison avec mitochondries
La présence de I' ADN permet de synthétiser une partie des protéines plastidiales (comme cela
se passe chez les procaryotes)
Les plastoribosomes synthétisent
- Une partie de certaines protéines du stroma (Ex, grosse sous unité de Rubisco) Quelques
protéines de structure des plastoribosomes
Une partie dès PSI, PSII, du cytochrome b6-f et de l'ATPsynthétase
Des facteurs d'élongation
3- ECHANGES ENTRE IIVALOPLASME et CHLOROPLASTES
Contrôlés par la membrane interne de l'enveloppe
-perméable aux petites molécules non chargées CO2, H2O, O2
- imperméable
aux ions chargés :H+, Mg++, Na+, K+.
Aux nucléotides (ADP ATP Pi, NADP),
Aux produits du cycle Caivin Bensson Bassham,
(Ils passent tous grâce à des transporteurs ou à des navettes actives le plus souvent).
1V -BIOGENESE
1) par division de chloroplastes préexistants (idem mitochondries et bactéries :
a- Partition : la membrane interne s’invagine perpendiculairement au grand axe et
partage le stroma en deux puis étranglementde la surface du chloroplaste.
b- Segmentation : étranglement de la région médiane.
2) par différenciation de proplastes
Le proplaste est un plaste indifférencié présent dans les cellules embryonnaitres végétales
(méristème, voir histologie végétale).
Lumière,- . .
Chloroplaste
(avec grana)
feuille verte
Proplaste
Lumière
Obscurité
Etioplaste
feuille blanchatre
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
1
/
26
100%