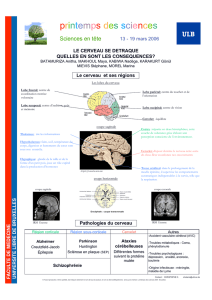
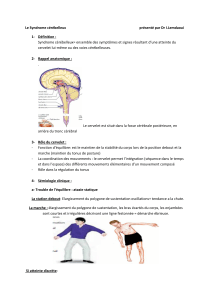
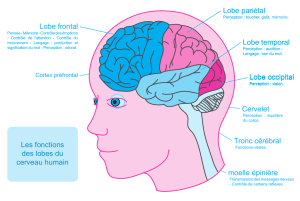
Organisation fonctionnelle du cervelet : approche neuro-anatomique
Telechargé par
ahmed salem Kleib

Revue ge
´ne
´rale
Organisation fonctionnelle du cervelet : approche
neuro-anatomique
Functional organisation of the cerebellum: A neuroanatomical approach
F. Vuillier
a,
*
,b
,E
´. Medeiros de Bustos
b
, L. Tatu
a,c
a
De
´partement d’anatomie, UFR sciences me
´dicales et pharmaceutiques, universite
´de Franche-Comte
´, 25000 Besanc¸on, France
b
Service de neurologie 2, ho
ˆpital Jean-Minjoz, CHU, 25000 Besanc¸on, France
c
Service d’explorations et pathologies neuro-musculaires, ho
ˆpital Jean-Minjoz, CHU, 25000 Besanc¸on, France
Les fonctions du cervelet ont initialement e
´te
´de
´duites des
travaux d’expe
´rimentation animale. Son implication dans les
fonctions motrices a e
´te
´confirme
´e plus tard chez l’homme a
`
travers de nombreux travaux cliniques aboutissant a
`la
description du syndrome ce
´re
´belleux (Thomas, 1897 ;
Babinski, 1899 ; Holmes, 1939). L’organisation cellulaire du
cervelet, assez proche de celle du cerveau, a elle e
´te
´de
´crite a
`
travers des travaux histologiques ou neurophysiologiques
(Ramon y Cajal, 1909 ; Eccles et al., 1967).
En recevant en permanence un ensemble d’affe
´rences
proprioceptives, vestibulaires et visuelles, le cervelet contri-
bue au maintien de l’e
´quilibre, a
`la re
´gulation du tonus
revue neurologique 167 (2011) 361–369
info article
Historique de l’article :
Rec¸u le 10 mars 2010
Rec¸u sous la forme re
´vise
´ele
3 mai 2010
Accepte
´le 20 juillet 2010
Disponible sur Internet le
6 mai 2011
Mots cle
´s:
Cervelet, Fonctions motrices,
Neuro-anatomie
Keywords:
Cerebellum, Motor functions,
Neuro-anatomy
re
´sume
´
La vision de l’organisation fonctionnelle du cervelet a e
´volue
´. Les divisions anatomiques et
phyloge
´ne
´tiques du cervelet qui ont longtemps servi de support a
`la description des
fonctions ce
´re
´belleuses sont de
´sormais trop re
´ductrices. La compre
´hension des fonctions
motrices du cervelet ne
´cessite la connaissance d’une division neuro-anatomique base
´e sur
les affe
´rences et l’organisation cortico-nucle
´aire du cervelet. Les fonctions non motrices du
cervelet sont encore mal syste
´matise
´es, mais les progre
`s rapides des neurosciences et
des techniques de neuro-imagerie fonctionnelle permettront certainement de mieux les
appre
´hender.
#2011 Elsevier Masson SAS. Tous droits re
´serve
´s.
abstract
Knowledge of the functional organisation of the cerebellum has progressed. The phyloge-
netic and anatomical divisions within the cerebellum, which have long been used to
describe cerebellar functions, are now too simplistic. The understanding of cerebellar motor
functions requires knowledge of a neuroanatomical division based on the afferents and
the corticonuclear organisation of the cerebellum. Non-motor cerebellar functions are
even more poorly systematised, but the rapid progress being made in neuroscience and
neuroimaging techniques should enable them to be better understood.
#2011 Elsevier Masson SAS. All rights reserved.
*Auteur correspondant.
Adresse e-mail : fabrice.vu[email protected] (F. Vuillier).
0035-3787/$ – see front matter #2011 Elsevier Masson SAS. Tous droits re
´serve
´s.
doi:10.1016/j.neurol.2010.07.045

musculaire, a
`la coordination de l’action des muscles au cours
du mouvement volontaire ainsi qu’a
`la programmation
motrice. Les travaux mene
´s plus re
´cemment ont cependant
fait sortir le cervelet de son roˆle exclusivement moteur. Il est
de
´sormais conside
´re
´comme un acteur intervenant dans le
domaine plus large de la cognition, des e
´motions ou du
langage (Leiner et al., 1986 ; Baillieux et al., 2008).
La vision de l’organisation fonctionnelle du cervelet a
e
´volue
´. Les divisions anatomiques et phyloge
´ne
´tiques du
cervelet qui ont longtemps servi de support a
`la description
des fonctions ce
´re
´belleuses sont de
´sormais trop re
´ductrices.
La compre
´hension des fonctions motrices ne
´cessite la
connaissance d’une division ce
´re
´belleuse base
´e sur les
affe
´rences et l’organisation cortico-nucle
´aire du cervelet.
Les fonctions non motrices ne sont pas encore syste
´matise
´es,
mais les progre
`s rapides des neurosciences et des techniques
de neuro-imagerie fonctionnelle permettront certainement de
mieux les appre
´hender.
Dans ce travail, apre
`s avoir rappele
´les grands aspects
morphologiques du cervelet et de son organisation cellulaire,
nous de
´crivons la division fonctionnelle du cervelet servant de
support a
`la compre
´hension de ces fonctions motrices. Les
principales donne
´es actuelles sur ses fonctions non motrices
sont ensuite pre
´sente
´es.
1. Aspects morphologiques
Le cervelet est une structure nerveuse appartenant a
`
l’ence
´phale et repre
´sentant pre
`s de 10 % du poids du cerveau
(Fig. 1 et 2). Il occupe la majeure partie de la fosse ce
´re
´brale
poste
´rieure en se plac¸ant en arrie
`re du tronc ce
´re
´bral auquel il
est rattache
´de manie
`re syme
´trique par les pe
´doncules
ce
´re
´belleux. Le cervelet est donc une structure situe
´een
de
´rivation du tronc ce
´re
´bral. Cet emplacement strate
´gique
explique ses modalite
´s d’action sur les grandes voies
nerveuses ascendantes et descendantes.
Le cervelet est classiquement de
´crit comme constitue
´de
trois parties : le vermis, impair et me
´dian, entoure
´de part et
d’autre par deux he
´misphe
`res pairs et syme
´triques. A
`sa
surface, le cervelet est parcouru par de nombreuses fissures et
sillons transversaux, plus ou moins profonds et concentri-
ques, qui contribuent a
`de
´limiter les lobules et les lobes.
La face supe
´rieure du cervelet est se
´pare
´edeshe
´misphe
`res
ce
´re
´braux par une structure dure-me
`rienne, la tente du
cervelet. Sa face infe
´rieure repose dans les fosses ce
´re
´belleuses
de l’os occipital. Sa face ante
´rieure est applique
´e contre la face
dorsale du tronc ce
´re
´bral constituant le toit du quatrie
`me
ventricule. Elle comporte les trois paires de pe
´doncules
ce
´re
´belleux, supe
´rieurs, moyens et infe
´rieurs, qui laissent
passer des faisceaux affe
´rents et effe
´rents unissant le cervelet
aux autres parties du syste
`me nerveux central.
2. Organisation cellulaire
Comme le cerveau, la configuration inte
´rieure du cervelet fait
apparaı
ˆtre de la substance grise corticale, ou cortex, de la
substance grise centrale, correspondant aux noyaux profonds
du cervelet et de la substance blanche.
2.1. Le cortex ce
´re
´belleux
Le cortex ce
´re
´belleux, tre
`s plisse
´, comporte trois couches
cellulaires diffe
´rentes dont la re
´partition est homoge
`ne : la
couche granulaire, la couche des cellules de Purkinje et la
couche mole
´culaire. Le cortex est organise
´fonctionnellement
autour de la cellule de Purkinje, dont l’axone constitue la seule
effe
´rence ce
´re
´belleuse.
La couche granulaire, la plus profonde, est une couche
re
´ceptrice qui contient essentiellement de nombreuses cellu-
les granulaires et des interneurones appele
´s cellules de Golgi.
La couche interme
´diaire est la couche effectrice du cortex,
forme
´e des cellules de Purkinje, les plus spe
´cifiques du
cervelet. Les axones des cellules de Purkinje gagnent les
noyaux gris du cervelet et les noyaux vestibulaires du tronc
ce
´re
´bral apre
`s avoir traverse
´la couche granulaire. La couche
mole
´culaire, la plus superficielle, est une couche associative
contenant des interneurones tels que les cellules a
`corbeille et
les cellules e
´toile
´es.
2.2. Substance blanche
La substance blanche du cervelet est forme
´e d’un ensemble
de fibres mye
´linise
´es affe
´rentes et effe
´rentes. Les fibres
affe
´rentes sont de deux types, les fibres grimpantes et les
fibres moussues. Les fibres grimpantes sont des axones en
[()TD$FIG]
fp
fs fl
Face
supérieure
Face
inférieure
Face
antérieure
Lobe antérieur
Lobe postérieur
Lobe flocculo-nodulaire
Fig. 1 – Coupe sagittale me
´diane du tronc ce
´re
´bral et du
cervelet. La face supe
´rieure du cervelet est en rapport avec
la tente dure-me
`rienne du cervelet. La face infe
´rieure
repose sur l’os occipital et la face ante
´rieure se projette en
regard du tronc ce
´re
´bral formant le toit du quatrie
`me
ventricule. Le lobe ante
´rieur est se
´pare
´du lobe poste
´rieur
par la fissure primaire (fp) et du lobe flocculo-nodulaire par
la fissure pe
´ote
´rolate
´rale (fl). Sur un plan fonctionnel, le
lobe ante
´rieur est plus spe
´cifiquement de
´volu aux
fonctions sensorimotrices et le lobe poste
´rieur aux
fonctions cognitives.
Median sagittal section of the cerebellum and brainstem.
(pf: primary fissure, sf: secondary fissure, plf: postero-lateral
fissure).
revue neurologique 167 (2011) 361–369362

provenance des olives bulbaires qui se ramifient pour s’accoler
aux dendrites des cellules de Purkinje. Les fibres moussues en
provenance du tronc ce
´re
´bral, en particulier des noyaux du
pont, et de la moelle spinale se terminent sur les dendrites des
cellules granulaires.
Les seules fibres effe
´rentes sont constitue
´es par les axones
des cellules de Purkinje qui projettent sur les noyaux
ce
´re
´belleux profonds et sur les noyaux vestibulaires.
2.3. Les noyaux ce
´re
´belleux profonds
Les noyaux ce
´re
´belleux profonds sont des amas, pairs et
syme
´triques, de substance grise intercale
´s dans la substance
blanche. Le noyau fastigial, le plus me
´dial, est situe
´dans la
substance blanche du vermis au contact du toit du quatrie
`me
ventricule. Les noyaux interpose
´s, regroupant les noyaux
globuleux et emboliforme, sont situe
´s dans la partie la plus
me
´diale de l’he
´misphe
`re. Les noyaux dente
´s, les plus
volumineux, sont place
´s plus late
´ralement dans l’he
´misphe
`re.
Les noyaux ce
´re
´belleux profonds rec¸oivent des affe
´rences
provenant du cortex ce
´re
´belleux par les axones des cellules de
Purkinje dispose
´es selon trois grandes zones longitudinales.
Les axones provenant du vermis se projettent sur le noyau
fastigial, ceux provenant de la zone interme
´diaire entre vermis
et he
´misphe
`re, se terminent sur les noyaux interpose
´s et enfin
ceux de la zone la plus late
´rale se projettent sur le noyau dente
´.
Ils ont tous une action inhibitrice sur les noyaux ce
´re
´belleux
profonds.
D’autres affe
´rences extrace
´re
´belleuses atteignent les
noyaux ce
´re
´belleux profonds par l’interme
´diaire de fibres
collate
´rales isssues des fibres grimpantes et des fibres
moussues. Elles exercent une action excitatrice sur les noyaux
ce
´re
´belleux profonds.
2.4. Circuits cellulaires
Les circuits cellulaires du cervelet sont e
´minemment comple-
xes et toujours sujets a
`discussion mais il est ne
´anmoins
possible de de
´limiter les grandes boucles de fonctionnement
(Eccles et al., 1967)(Fig. 3).
Les fibres moussues, en provenance du tronc ce
´re
´bral et de
la moelle spinale, s’articulent avec plusieurs cellules granu-
laires. Les fibres moussues ont une action excitatrice sur les
cellules granulaires et les interneurones de Golgi inhibent les
jonctions fibres moussues-cellules granulaires. Les articula-
tions synaptiques entre fibres moussues, cellules granulaires
et cellules de Golgi se de
´roulent au niveau de dispositifs
particuliers appele
´s glome
´rules.
[()TD$FIG]
m
s
i
V4
fp
fs
Efférences vers le noyau rouge
et le thalamus
Efférences vers la
formation réticulée
Afférences
cortico-pontiques
Afférences spinales,
vestibulaires et olivaires
Afférences spinales Efférences
spinales et vestibulaires
fl
f
Lobe antérieur
Lobe postérieur
Lobe flocculo-nodulaire
n
Fig. 2 – Vue ventrale du cervelet apre
`s ablation du tronc ce
´re
´bral. (n : nodule, f : floccule, fp : fissure primaire, fs : fissure
secondaire, fl : fissure poste
´rolate
´rale, V4 : quatrie
`me ventricule). Le pe
´doncule ce
´re
´belleux infe
´rieur (i) ve
´hiculent des
informations affe
´rentes en provenance essentiellement de la moelle spinale par le faisceau spinoce
´re
´belleux dorsal (de
Flechsig) qui rejoint le cervelet interme
´diaire (spinoce
´re
´bellum) ; du noyau cune
´iforme late
´ral (de von Monakow) par le
faisceau cune
´oce
´re
´belleux qui rejoint e
´galement le cervelet interme
´diaire (spinoce
´re
´bellum) et de l’olive bulbaire
(infe
´rieure) controlate
´rale, par le faisceau olivoce
´re
´belleux qui rejoint le cervelet late
´ral (ce
´re
´broce
´re
´bellum). Les fibres
effe
´rentes qui empruntent le pe
´doncule ce
´re
´belleux infe
´rieur gagnent la moelle spinale par le faisceau re
´ticulospinal et les
noyaux vestibulaires. Le pe
´donculde ce
´re
´belleux moyen (m) contient les fibres affe
´rentes en provenance des noyaux du
pont controlate
´raux se rendant au cervelet late
´ral (ce
´re
´broce
´re
´bellum). Le pe
´doncule ce
´re
´belleux supe
´rieur (s) permet le
passage des voies effe
´rentes du cervelet qui rejoignent le thalamus et le noyau rouge controlate
´raux apre
`s avoir croise
´la
ligne me
´diane au niveau du me
´sence
´phale (commissure de Wernekinck). Le faisceau spinoce
´re
´belleux ventral (de Gowers)
rejoint le cervelet interme
´diaire par le pe
´doncule ce
´re
´belleux supe
´rieur.
Ventral aspect of the cerebellum exposed by removal of the brainstem. (n: nodulus, f: flocculus, pf: primary fissure, sf: secondary
fissure, plf: postero-lateral fissure, V4: fourth ventricle, s: superior cerebellar peduncle, m: middle cerebellar peduncle, i: inferior
cerebellar peduncle).
revue neurologique 167 (2011) 361–369 363

Les fibres grimpantes, en provenance de l’olive bulbaire
controlate
´rale, se terminent par enroulement sur les dendrites
des cellules de Purkinje en contact avec les fibres paralle
`les.
Elles exercent une action excitatrice sur les cellules de
Purkinje. Les fibres grimpantes et les fibres moussues
e
´mettent des collate
´rales excitatrices pour les noyaux pro-
fonds du cervelet.
Par leurs longs axones, les cellules granulaires atteignent la
couche superficielle du cortex pour se bifurquer en T et former
les fibres paralle
`les qui stimulent les cellules de Purkinje. Les
fibres paralle
`les ont e
´galement une action excitatrice sur les
interneurones, cellules e
´toile
´es et cellules a
`corbeille, qui ont
elles-meˆmes une action inhibitrice sur la cellule de Purkinje.
Les axones des cellules de Purkinje constituent les seules
effe
´rences du cortex ce
´re
´belleux. Ils gagnent les noyaux
profonds du cervelet et les noyaux vestibulaires sur lesquels
ils exercent une inhibition.
3. E
´volution des concepts sur la division du
cervelet
La division initiale du cervelet e
´tait uniquement macros-
copique en un vermis me
´dian et deux he
´misphe
`res late
´raux.
Puis, le cervelet a e
´te
´parcellise
´en lobes par un syste
`me de
fissures. Ainsi, le lobe poste
´rieur est se
´pare
´du lobe ante
´rieur
par la fissure primaire et par la fissure secondaire. Le lobe
flocculo-nodulaire est se
´pare
´du lobe poste
´rieur par la fissure
poste
´rolate
´rale. Chacun de ces lobes est constitue
´d’une partie
me
´diane appartenant au vermis associe
´ea
`des extensions sur
les he
´misphe
`res (Fig. 1).
En conside
´rant le cervelet comme une structure anato-
mique ancienne dans l’e
´chelle animale et dont l’e
´volution est
diffe
´rente selon les espe
`ces, une division phyloge
´ne
´tique a
e
´galement e
´te
´propose
´e. Dans cette conception, le cervelet est
divise
´en trois parties correspondant a
`trois grandes e
´tapes
phyloge
´ne
´tiques (Larsell et Jansen, 1972). L’arche
´oce
´re
´bellum
est la partie la plus ancienne, propre aux verte
´bre
´s infe
´rieurs,
il correspond, en partie, au lobe flocculo-nodulaire. Le
pale
´oce
´re
´bellum est pre
´sent chez les reptiles et chez les
oiseaux. Il est constitue
´de la majeure partie du lobe ante
´rieur
et d’une petite partie du lobe poste
´rieur. Le ne
´oce
´re
´bellum est
propre aux mammife
`res et constitue la majeure partie du lobe
poste
´rieur.
Ne
´anmoins, la division la plus adapte
´ea
`l’organisation
fonctionnelle du cervelet et a
`la se
´miologie clinique est celle
base
´e sur l’origine des affe
´rences ce
´re
´belleuses et sur son
organisation cortico-nucle
´aire (Ito et al., 1990 ; Nieuwenhuys
et al., 1988). Cette division fonctionnelle est organise
´e selon le
plan sagittal et distingue les cervelets me
´dian, interme
´diaire
et late
´ral. Pour chacune de ces trois zones fonctionnelles et de
chaque coˆte
´, une partie du cortex est associe
´ea
`un noyau
profond.
Ainsi, le cervelet me
´dian correspond au cortex du vermis et
au noyau fastigial. Le cervelet interme
´diaire associe au cortex
de la partie me
´diale de l’he
´misphe
`re les noyaux interpose
´set
le cervelet late
´ral regroupe le cortex de la partie late
´rale de
l’he
´misphe
`re et le noyau dente
´. D’un point de vue fonctionnel,
le cervelet interme
´diaire et le cervelet late
´ral sont intimement
lie
´s. A
`ces trois zones fonctionnelles, s’ajoute le lobe flocculo-
nodulaire forme
´du nodule, structure appartenant au vermis,
et des ses prolongements late
´raux, les floccules. Classique-
ment, le noyau vestibulaire est conside
´re
´comme un ve
´ritable
noyau ce
´re
´belleux associe
´au lobe flocculo-nodulaire (Fig. 4).
Compte tenu de la pre
´ponde
´rance des affe
´rences spinales
des cervelets me
´dian et interme
´diaire, ils sont parfois appele
´s
spinoce
´re
´bellum. Le cervelet late
´ral est e
´galement appele
´
ce
´re
´broce
´re
´bellum, ses affe
´rences provenant en majeure
partie du cortex ce
´re
´bral. Enfin, le lobe flocculo-nodulaire
recevant principalement des affe
´rences vestibulaires est
e
´galement de
´fini comme le vestibuloce
´re
´bellum.
4. Fonctions motrices du cervelet
Au cours des deux derniers sie
`cles, les conceptions sur le
fonctionnement du cervelet ont beaucoup e
´volue
´. Les travaux
expe
´rimentaux pratique
´s chez l’animal ont permis d’envisa-
ger son roˆ le fonctionnel majeur dans les fonctions motrices.
Chez l’homme, les grandes descriptions cliniques neurologi-
ques du syndrome ce
´re
´belleux ont confirme
´son implication
dans le maintien de l’e
´quilibre, le re
´glage du tonus musculaire,
la coordination de l’action des muscles au cours du mouve-
ment volontaire et la programmation du mouvement.
[()TD$FIG]
Fig. 3 – Circuits cellulaires du cervelet. (1 : cellules
granulaires, 2 : cellules de Golgi, 3 : cellules e
´toile
´es, 4 :
cellules a
`corbeille, 5 : cellules de Purkinje). L’organisation
fonctionnelle du cortex ce
´re
´belleux est centre
´e sur la
cellule de Purkinje soumise a
`des influx excitateurs et
inhibiteurs. Les influx excitateurs proviennent des fibres
grimpantes et des fibres paralle
`les, elles-me
ˆmes contro
ˆle
´s
par les glome
´rules. Les effets inhibiteurs sont assure
´s par
les cellules e
´toile
´es et les cellules a
`corbeille. Cette double
influence sur la cellule de Purkinje est a
`l’origine de
potentiels d’action dont la seule voie de sortie est
repre
´sente
´e par son axone qui projettent vers les noyaux
ce
´re
´belleux et les noyaux vestibulaires.
Cellular cerebellar circuitry. (1: mossy fibers, 2: Golgi cells
granule cells, 3: stellate cells, 4: basket cells Golgi cells, 5:
Purkinje cells).
revue neurologique 167 (2011) 361–369364

4.1. Re
´gulation du mouvement volontaire des membres –
cervelet interme
´diaire
La fonction principale du cervelet interme
´diaire est la
coordination de l’action des muscles distaux au cours du
mouvement volontaire des membres (Ito, 1984 ; Nieuwenhuys
et al., 1988 ; Duvernoy, 1995)(Fig. 5). Le cervelet interme
´diaire
rec¸oit essentiellement des affe
´rences homolate
´rales prove-
nant de la pe
´riphe
´rie. Ces affe
´rences sont ve
´hicule
´es par les
faisceaux spinoce
´re
´belleux, cune
´oce
´re
´belleux et trige
´mino-
ce
´re
´belleux. Les faisceaux spinoce
´re
´belleux correspondent a
`
la sensibilite
´proprioceptive inconsciente. Le faisceau cune
´o-
ce
´re
´belleux constitue la partie la plus late
´rale de la voie
proprioceptive consciente ve
´hiculant des informations
appartenant a
`la sensibilite
´e
´picritique. Ce faisceau fait relais
dans le noyau cune
´iforme late
´ral du bulbe (noyau de von
Monakow) avant d’atteindre le cervelet. Enfin, le faisceau
trige
´minoce
´re
´belleux ame
`ne les informations proprioceptives
inconscientes en provenance de la face.
Ces faisceaux se projettent sur le cortex interme
´diaire de
l’he
´misphe
`re et fournissent au cervelet des informations sur
l’exe
´cution du mouvement en cours. Par l’interme
´diaire des
noyaux interpose
´s, ses principales effe
´rences envoient des
messages au noyau ventral late
´ral du thalamus controlate
´ral
et ensuite au cortex moteur.
Par cette voie, le cervelet peut alors moduler le fonction-
nement de la voie motrice principale. Il re
´ajuste les parame
`-
tres cine
´matiques (dure
´e, direction, vitesse et amplitude des
mouvements distaux) en particulier ceux de la main. Au de
´but
du mouvement, le cervelet interme
´diaire a une action
excitatrice sur les muscles agonistes pour ensuite exercer
une action excitatrice sur les muscles antagonistes. Cette
seconde action permet de freiner le mouvement et d’atteindre
le but avec pre
´cision. Par la meˆme, il intervient aussi dans
l’arreˆt du mouvement.
Depuis les noyaux interpose
´s, des effe
´rences gagnent aussi
la substance re
´ticule
´e, origine du faisceau re
´ticulospinal, dont
un des roˆles serait de moduler le tonus musculaire. Moins
importantes chez l’homme, des effe
´rences gagnent aussi le
paleorubrum, la partie magnocellulaire du noyau rouge, point
de de
´part de la voie rubrospinale dont le roˆ le serait d’aider a
`la
re
´alisation pre
´cise du geste final.
4.2. Programmation du mouvement – cervelet late
´ral
Durant le de
´roulement du mouvement en cours, la fonction
principale du cervelet late
´ral est la programmation du
mouvement suivant. Il intervient e
´galement dans l’apprentis-
sage de nouveaux mouvements (Fig. 6). Son roˆle est donc
d’assurer l’enchaı
ˆnement progressif et coordonne
´des mouve-
ments successifs (Nieuwenhuys et al., 1988 ; Duvernoy, 1995).
De manie
`re simplifie
´e, on peut conside
´rer que le cortex
moteur envoie au cervelet late
´ral controlate
´ral une copie du
mouvement pre
´vu. Par ses effe
´rences, le cervelet late
´ral
module alors le cortex moteur par une boucle ferme
´e
caracte
´rise
´e par un double de
´cussation. Le roˆle du cervelet
late
´ral est donc de programmer au mieux le mouvement
suivant et de limiter l’action correctrice effectue
´e en temps
re
´el par le cervelet interme
´diaire.
Le cervelet late
´ral ne rec¸oit pas d’affe
´rences provenant de
la pe
´riphe
´rie. Il rec¸oit essentiellement des fibres provenant du
cortex moteur controlate
´ral qui, avant de de
´cusser, font relais
dans les noyaux du pont. Le cortex ce
´re
´belleux projette sur le
noyau dente
´dont les effe
´rences, apre
`s avoir de
´cusser, se
dirigent vers le noyau ventral late
´ral du thalamus controla-
te
´ral avant de revenir au cortex moteur.
Un circuit secondaire centre
´sur l’olive bulbaire vient se
greffer sur cette boucle fonctionnelle principale cortico-ponto-
ce
´re
´bello-thalamo-corticale. L’olive bulbaire rec¸oit des infor-
mations e
´manant du cortex moteur homolate
´ral soit directe-
ment, soit apre
`s relais dans le ne
´orubrum, la partie
parvocellulaire du noyau rouge. Par le faisceau spino-olivaire,
l’olive bulbaire rec¸oit e
´galement des informations pe
´riphe
´ri-
ques provenant de la moelle spinale. De l’olive bulbaire
partent des effe
´rences stimulatrices vers le cervelet late
´ral
oppose
´.A
`long terme ces effe
´rences jouent un roˆ le important
dans l’apprentissage moteur.
[()TD$FIG]
LATERAL INTERMEDIAIRE
MEDIAN
Noyau denté Noyaux interposés
Noyaux
fastigiaux
LOBE FLOCCULO-NODULAIRE
d
i
f
Afférences spinales (axe du corps et tronc)
Afférences spinales (membres)
Afférences corticales
Afférences vestibulaires et visuelles
N
F
F
d
i
f
Fig. 4 – Divisions fonctionnelles du cervelet en cervelets
me
´dian, interme
´diaire, late
´ral et lobe flocculo-nodulaire.
Le cervelet interme
´diaire est forme
´du cortex de la partie
me
´diale de l’he
´misphe
`re et des noyaux interpose
´s (i). Il
contro
ˆle les mouvements volontaires des membres. Le
cervelet me
´dian associe le cortex du vermis et le noyau
fastigial (f) pour assurer le contro
ˆle des muscles axiaux et
des muscles proximaux des membres. Du fait de leurs
affe
´rences essentiellement spinales, les cervelet me
´dian et
interme
´diaire sont regroupe
´s sous le terme de
spinoce
´re
´bellum. Le cervelet late
´ral (ce
´re
´broce
´re
´bellum)
qui regroupe le cortex de la partie late
´rale de l’he
´misphe
`re
ce
´re
´belleux et le noyau dente
´(d) rec¸oit essentiellement
des affe
´rences corticales. Sa fonction est la
programmation du mouvement. Le lobe flocculo-nodulaire
(vestibuloce
´re
´bellum) regroupe le nodule (N) et les
floccules (F). Il rec¸oit essentiellement des affe
´rences
vestibulaires et assure le contro
ˆle de l’e
´quilibre axial.
Functional division of the cerebellum: medial cerebellum,
intermediate cerebellum, lateral cerebellum and flocculo-
nodulus lobule. (f: fastigial nucleus i: interposed nuclei,
d: dentate nucleus).
revue neurologique 167 (2011) 361–369 365
 6
7
8
9
6
7
8
9
1
/
9
100%