Lycée Regnault Ibtisam Boudouaia
1
2017-2018
Chapitre II : la dynamique du génome
Notions : Des anomalies peuvent survenir. Un crossing-over inégal aboutit parfois à
une duplication de gène. Un mouvement anormal de chromosomes produit une cellule
présentant un nombre inhabituel de chromosomes. Ces mécanismes, souvent
sources de troubles, sont aussi parfois sources de diversification du vivant (par
exemple à l'origine des familles multigéniques).
Notion : D'autres mécanismes de diversification des génomes existent : hybridations
suivies de polyploïdisation, transfert par voie virale, etc.
Rappel des acquis :
En seconde : À l’échelle d’une espèce, la biodiversité correspond à la diversité des
allèles des différents gènes et à leur fréquence dans les différentes populations de
cette espèce.
En première : les mutations géniques créaient de nouveaux allèles. Au niveau d’une
espèce, la biodiversité génétique est donc le résultat d’une accumulation de ces
mutations.
Le chapitre précédent a permis de comprendre que la reproduction sexuée par les
mécanismes de la méiose et de la fécondation permettait de créer de nouvelles
combinaisons d’allèles d’une génération à l’autre.
Pourtant, l’association de mutations et du brassage génétique au cours de la méiose puis de
la fécondation ne suffit pas à expliquer la diversification génétique des êtres vivants.
Pb : Quels sont les processus autres que les mutations et le brassage génétique
pouvant rendre compte de la complexité des génomes et donc de la diversification
génétique des êtres vivants ?
I- Les modifications du nombre de chromosomes
A- La non ségrégation des chromosomes ou des chromatides lors de la
méiose
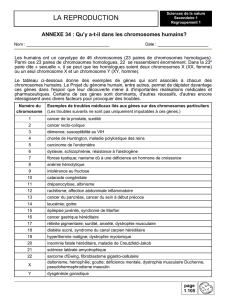
Etude de caryotypes d’individus atteints d’aneuploïdie (anomalies du nombre de
chromosomes)

Lycée Regnault Ibtisam Boudouaia
2
2017-2018
Thème 1-A-2 Diversification génétique et diversification des êtres vivants TS OBLI
ACTIVITE 1 : Etude de quelques anomalies du nombre de
chromosome
Eléments de correction pour l’ACTIVITE 1 : Etude de quelques anomalies du nombre
de chromosome
La perturbation responsable de la formation d´un gamète contenant deux chromosomes 21
peut survenir durant la première ou la deuxième division de la méiose.
Durant la première division : la non-séparation des
chromosomes homologues :
Les deux chromosomes 21, toujours formés de deux
chromatides, migrent du même côté de la cellule lors
de l´anaphase I.
Une des deux cellules à l´issue de la télophase I
possède alors deux chromosomes 21 et l´autre aucun.
La deuxième division aboutit à la formation de deux
cellules possédant deux chromosomes 21 simples.
Durant la deuxième division : la non-séparation
des chromatides :
La première division se déroule normalement et
donne naissance à deux cellules filles qui
contiennent chacune un chromosome 21 formé de
deux chromatides. Lors de la deuxième division,
les deux chromatides se séparent mais migrent
vers le même pôle de la cellule donnant ainsi
naissance à un gamète possédant deux
chromosomes 21.
Ensuite à la fécondation, il y a fécondation d´un ovocyte contenant deux chromosomes 21
par un spermatozoïde n´en contenant qu´un seul. Cela donne une cellule œuf d´individu
trisomique.
Schéma pour la translocation robertsionienne

Lycée Regnault Ibtisam Boudouaia
3
2017-2018
Les patients porteurs d’une translocation robertsonienne ont un caryotype à 45
chromosomes. Le fragment composé des bras courts des acrocentriques est perdu. Lors de
la méiose, Il existe un risque de formation de gamètes déséquilibrés donnant des zygotes
trisomiques ou monosomiques.
Conclusion :
Les anomalies de nombre peuvent se traduire par l’absence d’un ou de plusieurs
chromosomes ou la présence d’un chromosome supplémentaire dans le caryotype de
l’espèce.
Les anomalies de nombre s’expliquent le plus souvent par la non-disjonction des
chromosomes homologues au cours de l’anaphase de la première division de méiose ou par
la non-disjonction des chromatides lors de la seconde phase de la méiose.
Au final, deux chromosomes d’une même paire au lieu d’un seul peuvent alors se retrouver
dans un même gamète. Si ce gamète est fécondé par un gamète normal, l’œuf qui en
résultera sera donc trisomique : si le gamète ne contient aucun chromosome de la paire
considérée, l’œuf sera monosémique.
B- Polyploïdie etdiversification des génomes
Indication : La ploïdie est le nombre de jeux de chromosomes, c’est à dire le nombre de
copies des différents chromosomes formant le génome. Dans le noyau des cellules
humaines, il y a deux copies de chaque chromosome. On parle alors de diploïdie. Quand le
nombre de lots de chromosomes est supérieur a` deux, on parle de polyploïdie.

Lycée Regnault Ibtisam Boudouaia
4
2017-2018
Thème 1-A-2 Diversification génétique et diversification des êtres vivants TS OBLI
ACTIVITE 2 : Polyploïdie et diversification des génomes
Eléments de correction : ACTIVITE 2 : Polyploïdie et diversification des génomes
Correction :
La figure de métaphase d’une mitose Triticum monococcum montre distinctement 14
chromosomes ; celle de métaphase de méiose 1 montre 7 paires de chromosomes
appariés. Cela conduit à la formule chromosomique 2n=14. Il s’agit donc du
caryotype d’une espèce diploïde.
La figure de métaphase de mitose de Triticumturgidum (durum) montre 28
chromosomes et celle de méiose 1 montre 14 paires de chromosomes appariés. On
arrive dont à la formule chromosomique 2n=28.
Enfin, la photo de métaphase de mitose de Triticumaestivum permet plus difficilement
de compter le nombre de chromosomes : 42 mais sur la photo de méiose 1 on
distingue nettement 21 paires de chromosomes appariés. La formule chromosomique
est donc 2n=42 chromosomes.
Avec ces documents, on ne peut conclure à la polyploïdie des blés dur et tendre car le
comportement des chromosomes à la méiose chez ces deux espèces est typique de celui
d’une espèce diploïde.
On peut seulement constater que les trois espèces de blé diffèrent par leur nombre de
chromosomes et que ce nombre est un multiple de 7.
Est ce que cela est dû au hasard ou à un mécanisme à l’œuvre lors de la formation des deux
espèces de blé dur et tendre ?
Comparaison des caryotypes
Le caryotype de Triticum monococcum présente toutes les caractéristiques d’un caryotype
classique. Il s’agit donc du caryotype d’une espèce diploïde.
En revanche les caryotypes du blé dur et du blé tendre présentent une originalité.
Celui du blé dur montre deux groupes de 7 chromosomes, désignés par les lettres A et B.
Dans chaque groupe, chaque chromosome est représenté en deux exemplaires ce qui est
habituel (chromosomes homologues).
Le caryotype du blé tendre montre, outre les deux groupes de chromosomes A et B, un
troisième groupe de 7 chromosomes, D.
Chez une espèce comme Triticum turgidum où le nombre de chromosomes est de 28, les
chromosomes du caryotype devraient être numérotés de 1 à 14 et chez le blé tendre (42
chromosomes) de 1 à 21. Ce n’est pas le cas.

Lycée Regnault Ibtisam Boudouaia
5
2017-2018
On identifie seulement 7 chromosomes ayant chacun un représentant dans les groupes A et
B (A1 et B1, A2 et B2, etc.) chez le blé dur, et un représentant dans les groupes A, B et D
chez le blé tendre.
Ainsi, on voit que le Triticum turgidum a 4 exemplaires de chaque chromosome et est donc
tétraploïde ; Triticum aestivum a 6 exemplaires de chaque chromosome et est donc
hexaploïde.
TOP :http://lewebpedagogique.com/tleclerc/files/2016/10/activite-GE6v1-ble-corrige.pdf
Comparaison des séquences de GLU
La comparaison des séquences nucléotidiques des gènes GLU (gènes de gluténines) des
lots A,B et C du blé tendre à l’aide d’anagène montre :
• 72,3% d’identité entre GLU-A et GLU-B
• 77% d’identité entre GLU-A et GLU-D
• 73,3% d’identité entre GLU-B et GLU-D
Ce fort degré d’homologie confirme que les trois séquences ont une origine commune : elles
sont issues d’un unique gène GLU ancestral présent chez l’espèce ancestrale à partir de
laquelle la famille des blés s’est diversifiée.
Remarque : ce degré d’homologie est cependant moins élevé que celui qui existe entre des
allèles du même gène. En effet, ces gènes différents ont évolué indépendamment les uns
des autres au fil des générations et accumulé des mutations différentes
Les gènes GLU-A, GLU-B et GLU-D sont homéologues.
Comparaison des séquences des gènes GLU de T. monococcum, T. speltoides et T. Tauchii
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%