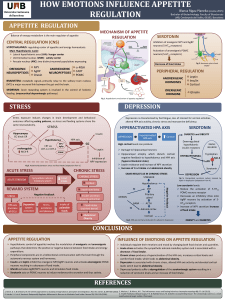
Ž.
Brain Research Reviews 32 2000 16–28 www.elsevier.comrlocaterbres
Short review
ž/
Electrical synapses, a personal perspective or history
Michael V.L. Bennett )
Department of Neuroscience, Albert Einstein College of Medicine, 1300 Morris Park AÕenue, Bronx, NY 10461, USA
Abstract
Gap junctions are the morphological substrate of one class of electrical synapse. This memoir records the author’s involvement in the
development of our knowledge of the physiology and ultrastructure of electrical synapses. The answer to whether neurotransmission is
electrical or chemical is either. One lesson is that Occam’s razor sometimes cut too deep; the nervous system does its operations in a
w
number of different ways and a unitarian approach can lead one astray M.V.L. Bennett, Nicked by Occam’s razor: unitarianism in the
Ž. x
investigation of synaptic transmission, Biol. Bull. 168 1985 159–167 . Electrical synapses can do many things that chemical synapses
can do, and do them just as slowly. The new molecular, cellular and physiological techniques will clarify where gap junctions and
electrical coupling do and do not occur and permit experimental manipulation with high specificity. q2000 Elsevier Science B.V. All
rights reserved.
Keywords: Gap junction; Electrical synapse; Connexin; Coupling
Contents
1. Introduction ........................................................................ 16
2. An aside on ‘‘ephapses’’ and other nomenclatural niceties ............................................... 17
3. The first connexin based electrical synapses! ...................................................... 18
4. What next? Electric organ control systems ....................................................... 20
5. Fast motor systems .................................................................... 22
6. Electrical versus chemical ................................................................ 24
7. An aside about evolution................................................................. 24
8. Romance in academia .................................................................. 25
Acknowledgements...................................................................... 27
References .......................................................................... 27
1. Introduction
It seems like yesterday that I started to work on electri-
cal communication between neurons. Well, actually not, it
seems quite a long time ago, and the field, and I, have
matured significantly over the years. The operation of
neurons at the level of electrical signaling, at least at the
level of questions that most of us were asking more than
)Tel.: q1-718-430-2536; fax: q1-718-430-8944; e-mail: [email protected]
0165-0173r00r$ - see front matter q2000 Elsevier Science B.V. All rights reserved.
Ž.
PII: S0165-0173 99 00065-X

()
M.V.L. BennettrBrain Research ReÕiews 32 2000 16–28 17
40 years ago, is pretty well understood, not completely of
course, but there are no very large black boxes. It seems
much more like yesterday, and is, that I last wrote a review
on the development of knowledge of electrical transmis-
wx
sion 73 . What has happened in the last couple of years?
Aside from substantial but incremental progress, there has
wx
been the discovery of neuron specific connexins 26,63
and of connexin diseases that involve the nervous system
wx
15,22,38 . Although no diseases of electrical synapses
have been discovered yet, there are sure to be mutations of
neuron specific connexins, whether or not the mutations
lead to an observable phenotype. Interestingly, mutations
of Cx26 and Cx32, which are expressed by some neurons
in the CNS, have yet to show any phenotypic changes in
those neurons. Gene targeting is progressing through the
connexin gene family. And there is the discovery that gap
Ž.
junctions of the Ecdysozoa the nematode, arthropod line
are encoded by a gene family completely unrelated to the
Ž
connexins, although there are many convergent not con-
.
served properties between the junctions encoded by that
gene family and by the connexins. Since the new connex-
ins and connexin diseases are extensively discussed in this
volume by others more directly involved in the research,
and I just participated in writing a review of connexin
wx
diseases 15 , I will make this presentation more an oral
history than an integrative summary and analysis.
One does not have to have been at Oxford to define a
synapse as a specialized site of functional interaction
Ž.
between neurons, although Sherrington and Eccles and I
were. By this definition gap junctions form one class of
wx
electrical synapse 13 . Another kind of electrical synapse
mediates short latency inhibition of the Mauthner cell of
teleost fishes and possibly mammalian cerebellar Purkinje
cells; this form of electrical transmission is not mediated
by gap junctions, and involves different junctional special-
Ž.wx
izations see below 28 . In addition, there probably are
electrical interactions that occur between closely apposed
cells without obvious gap junctions or specializations other
wx
than the absence of interposed glia 28,36,67 . Whether
these sites are to be considered synapses, i.e., specialized,
or ephapses, i.e., incidental or accidental sites of interac-
tion, may become clear with greater knowledge of the
developmental mechanisms. Without deciding on a name
one can still describe the electrical interaction, which does
indeed appear to be uniquely associated with the close
appositions.
2. An aside on ‘‘ephapses’’ and other nomenclatural
niceties
wx
Angelique Arvanitaki 1 coined the term ephapse from
the Greek to mean an apposition that is not quite so close
Ž
as a synapse according to my Greek–American colleague,
.
George Dimitrios Pappas . She used it to denote what she
thought of as artificial synapses, which she made by laying
one axon along side another where the ‘‘action currents’’
generated during an impulse in one axon altered the ex-
citability of the other axon. This terminology suggests that
she thought synaptic transmission was electrical, at least
that is my recollection, and, in the spirit of oral history, I
will not go to the library to try to confirm that view.
Ephapse then came to be used for incidental contacts in the
nervous system, particularly where activity in one or more
axons excited other axons. These days one might wonder if
actual gap junction electrical synapses were formed be-
tween axons in injured tissue.
My mentor at that time, Harry Grundfest, had embraced
the idea that chemical transmission was mediated at elec-
trically inexcitable membrane, i.e., in explicit, modern
terms that the conductance of the neurotransmitter recep-
tors was independent of membrane potential and only a
function of transmitter concentration. To keep transmission
at synapses chemically pure, he decided to use ephapse to
denote morphological specializations between neurons
where transmission was electrical. Although my Oxford
education led me to disagree with this practice, to keep
peace in the laboratory I used ‘‘electrotonic junctions’’ for
one type of what I now freely term electrical synapses. At
that time I felt that ephapse connoted artificiality or inci-
dentality, which downgraded the importance of my work.
Electron microscopy now makes it clear that gap junctions
are closer appositions than occur at chemical synapses,
and, if one were starting over, one would call chemical
synapses ephapses and gap junctions between neurons
Ž.
synapses. In my view both he and Jack later Sir John
Eccles made a mistake in thinking that only one mode of
transmission could be synaptic, but Eccles was better at
wx
changing his mind 14 .
I recall when David Potter presented his and Edward
wx
Furshpan’s work 31 on the crayfish giant motor synapse
at the Monday night electrobiology seminars at the Marine
Ž.
Biological Laboratory MBL, Woods Hole . This work and
wx
the independent studies of Akira Watanabe 70 on the
cardiac ganglion of the mantid shrimp were the first
unequivocal demonstrations of electrical transmission be-
tween neurons. The electrobiology sessions were organized
by Harry, who welcomed airing of all views, and were
known as the Monday Night Fights because of the some-
times heated discussion and by analogy with the Friday
Night Fights, a popular program at the time showing
professional boxing matches. Harry suggested in the dis-
cussion period, or possibly before, since interruptions were
not uncommon, that they should call their rectifying
synapse an ephapse, because it was electrical although it
was electrically inexcitable. David pointed out to him with
evident pleasure that the junctional conductance was a
function of voltage and thus was electrically excitable.
Harry did not have a good retort, which was unusual for
him. Of course gap junctions between segments of the
septate axon are electrically linear over a wide range
wx
37,71 , while connexin based gap junctions all show some

()
M.V.L. BennettrBrain Research ReÕiews 32 2000 16–2818
Ž
degree of dependence on transjunctional voltage e.g., Ref.
wx.
34 .
In a collection of gap junction papers, it should not be
necessary to obsess about the relative merits of chemical
and electrical synapses. We workers in the field for the
most part think highly of what we do. Still, in this contri-
bution it may be worthwhile to do a little complaining,
qvetching or whining, depending on one’s ethnic origin.
For example, consider the ‘‘medical subject headings’’
Ž.
MeSH for the PubMed data base. Under ‘‘Synapses’’ is:
Specialized junctions at which a neuron communicates
with a target cell. At classical synapses, a neuron’s
presynaptic terminal releases a chemical transmitter
stored in synaptic vesicles which diffuses across a
narrow synaptic cleft and activates receptors on the
postsynaptic membrane of the target cell. The target
may be a dendrite, cell body, or axon of another neuron,
or a specialized region of a muscle or secretory cell.
Neurons may also communicate through direct electri-
cal connections which are sometimes called electrical
synapses; these are not included here but rather in GAP
JUNCTIONS.
And then under ‘‘Synaptic Transmission’’:
Ž
The communication from a neuron to a target neuron,
.
muscle, or secretory cell across a synapse. In chemical
synaptic transmission, the presynaptic neuron releases a
neurotransmitter that diffuses across the synaptic cleft
and binds to specific synaptic receptors. These activated
receptors modulate ion channels andror second-mes-
senger systems to influence the postsynaptic cell. Elec-
trical transmission is less common in the nervous sys-
tem, and, as in other tissues, is mediated by gap junc-
tions.
Thus, to look for the latest in electrical transmission or
electrical synapses, one has to take a somewhat devious
route and examine all those other citations that come along
with gap junctions and nervous system. Looking for ear-
lier papers is more complicated because Gap Junctions as a
MeSH term was not introduced until 1994, and you cannot
search for the phrase ‘‘electrical synapse’’, although ‘‘elec-
trically synaptic transmission’’ is in the Compound Word
Dictionary and yields 12 citations. Another bit of whining
for the in-group: even some workers in the gap junction
field have trouble using electrical with respect to PSPs, no
doubt influenced by Harry. Korn and Faber write about
‘‘coupling potentials’’ at electrical synapses rather than
wx
PSPs. J.G.R. Jefferys 36 in a Physiological Review con-
siders ‘‘four classes of non-synaptic interaction, mainly in
the mammalian brain’’ of which the first is ‘‘Electrotonic
Ž.
and chemical coupling through gap junctions’’. Yet he
also writes of ‘‘gap junctions, which commonly serve as
electrical synapses in invertebrates but appear to be used
less often for electrical signaling in vertebrates’’.
The reader may feel that there is too much discussion of
terminology here. There probably was too much quar-
relling over nomenclature, but some of the controversy
represented real differences in concepts rather than ego-
driven preference. We should all be familiar with Feld-
berg’s Dictum, which is that a scientist would rather use
another scientist’s toothbrush than his terminology. I be-
Ž.
lieve I heard this from Sir Bernard Katz, who cited it in
one of his lectures. It is a delightfully apt phrase in that
words have a flavor of their origin andror meaning. When
I was a child, a not uncommon punishment for use of foul
language was washing the offender’s mouth out with soap;
thus, the mystical view seems to be that dirty words
physically soil the speaking apparatus. In my own case,
speaking of alpha and beta connexins make me want to
brush my teeth with my toothbrush.
Jean Paul Changeaux once chided me for calling gap
junctions between neurons electrical synapses, when the
same structures were called gap junctions when they oc-
curred between non-neuronal cells. The venue was a side-
walk cafe in Paris, and the statement should not be taken
very seriously. Moreover, as chemical interactions between
non-neuronal cells have become more widely described
and as the molecules responsible for exocytosis and endo-
cytosis at synapses have proved to have homologs in
non-neuronal cells, the same criticism might be lodged
about the terminology for chemical synapses.
3. The first connexin based electrical synapses!
The supramedullary neurons of the puffer fish,
Spheroides maculatus, were the subject of my first experi-
ments at the MBL in Woods Hole. These large neurons
Ž.
0.2–0.3 mm in diameter sit on the dorsal surface of the
medulla and can be seen with the naked eye, at least with
Ž.
my eyes at that time Fig. 1 . I no longer remember where
Harry Grundfest found out about them, possibly from
Shigehiro Nakajima and Susumu Hagiwara, who later
studied their action potential generation, but the cells were
known to early comparative anatomists including Sigmund
Freud. Large neurons were of interest, because they were
relatively easy to study with sharp intracellular microelec-
trodes and patch electrodes were far in the future. The
function of the neurons was unknown, and we were able to
show that they were effector cells sending their axons out
the dorsal roots to the skin. But nothing obvious happened
in the skin when they were stimulated. Recent data demon-
strate that they contain gastrinrcholecystokinin and inner-
wx
vate mucous glands 29 , and a secretomotor function
should be more carefully investigated. But that is compara-
tive physiology, which is primarily of interest to NSF.
What is more relevant to general physiology and this

()
M.V.L. BennettrBrain Research ReÕiews 32 2000 16–28 19
Fig. 1. Anterior spinal cord of the puffer viewed from the dorsal side. The posterior limit of the cerebellum is to the left. The supramedullary neurons are
the large round cells, about 250 mm in diameter, that are located on the surface of the cord. Several of the cells that had been penetrated for intracellular
wwxx
recording are dark because of increased staining by toluidine blue applied to the surface from Ref. 19 .
discussion is the observation that they fire synchronously
in response to cutaneous inputs.
The initial observation of synchronous firing was the
accidental result of putting two electrodes in adjacent cells
when trying to get two into one cell for separate current
application and voltage recording. What was one to think
in 1957 when one saw synchronous firing? I thought that a
higher level synchronizing center was exciting the
supramedullary neurons, and that the large depolarization
Ž
that initiated the overshooting spike was a PSP implicitly
.
chemically mediated generated by inputs from that center.
One afternoon Eccles was visiting the laboratory while I
was recording. He looked at the oscilloscope screen, saw
the two component spike and said of my synchronizing
input ‘‘That’s an initial segment spike’’. He advised me to
advance an electrode beneath the cluster of cell bodies to
record from the axons directly. I tried it, and, of course, he
was right. It was quite easy to find two component spikes
that characterized an axonal recording and then to hyper-
polarize one by one the overlying somata until the soma of
origin was identified. But in addition to finding axon
spikes and clarifying the nature of the two components of
the spike recorded in the soma, I also found coupling. The
cells are coupled by gap junctions between their axons
wx
20 . Thus, when one hyperpolarized an overlying cell
body that did not belong to an axon being recorded from,
the hyperpolarization due to coupling was larger than in
the cell body giving rise to that axon and occasionally big
enough that even the unprepared mind could not miss it.
We were rapidly convinced that the coupling was responsi-
ble for the synchronization. What had not been obvious is
that mutual excitation between the cells was electrical. An
action potential directly evoked in one cell could spread to
the rest of the cells in the cluster and this spread showed
paired pulse facilitation, although we did not call it that.
The period of increased excitability could be as long as
200 ms, which we thought suggested a chemical rather
than electrical mechanism, but subsequently it proved to be
explained by a long-lasting depolarizing afterpotential. And
the rest is history.
Let us be frank here. The presence of coupling was put
in a footnote in the 1959 puffer papers and discussed at
slightly greater length in a few abstracts. Full publication
wx
took about 7 years 9,20 . NIH was more forgiving and the
race for priority was not so hectic as it is now. Nor did we
know that connexins and Ecdysozoan gap junction proteins
were different families.
Although many of the implications of electrical cou-
pling of supramedullary neurons were not immediately
obvious and only became clear as other examples of
coupling were discovered, the system has relevance to
mammalian systems. First, electrical synapses can serve a
synchronizing function, but the degree of synchronization
need not be very precise; the spikes in different cells in the
cluster can be quite dispersed in time. Propagation of
impulses between cells can be slow compared to the delays
at chemical synapses and in some species the safety factor
for propagation can be less than one in that an impulse in

()
M.V.L. BennettrBrain Research ReÕiews 32 2000 16–2820
one cell is not always accompanied by an impulse in all
the other cells; the safety factor depends on the presynaptic
action potential, strength of coupling and excitability of the
postsynaptic cell. These conclusions could also have been
wx
drawn from Watanabe’s mantid shrimp data 70 . He ob-
served coupling of bursting neurons controlling heart rate.
He thought the cells were connected by cytoplasmic
bridges, which is probably wrong, but recognized the
synchronizing function of the coupling. Second, the syn-
chronizing function of electrical synapses involves hyper-
polarization of more positive cells, as well as depolariza-
tion of more negative cells. Coupling is a two way street,
and a significant fraction of a cell’s input conductance can
be the input conductance into its electrical synapses with
other cells. Third, impulses are more likely to spread
between cells when they are depolarized by other synaptic
inputs, chemical as well as electrical, and there can be both
spatial and temporal summation of electrical PSPs. All
these features are trivial if not obvious consequences of
coupling by gap junctions. Although we were concerned
about the morphological basis of electrical coupling, the
puffer was not a great preparation in which to look for gap
junctions, and associating gap junctions with electrical
transmission came later.
4. What next? Electric organ control systems
This was not the time when hunting for further electri-
cal synapses crossed my mind, but it proved possible to
blunder upon them. Harry Grundfest had been interested
Ž
before I arrived at P&S the College of Physicians and
.
Surgeons of Columbia University in how weakly electric
fishes generated their electric pulses and how the organ
discharge was controlled. These fishes, depending on the
species, emit brief pulses with relatively long intervals
between them or pulses that are separated by an interval
Ž.
comparable to the pulse duration Fig. 2 . The former
group, pulse fish, modulated their discharge frequency in
response almost any mode of stimulation, whereas the
latter group, wave fish, tended to have a very constant
frequency. Stimulating the spinal cord of wave fish did not
cause acceleration. When examined carefully, there was a
slight phase advance, which we now know to be due to
depolarization from antidromic activity spreading into the
pacemaker nucleus in which the frequency is set. Still, the
constancy was impressive in the face of a stimulus that
activated sensory inputs and caused a dramatic accelera-
tion in the pulse fish. Akira Watanabe, he who had shown
the coupling between cardiac ganglion cells of the mantid
shrimp, took a more subtle approach. Asking what a wave
fish would do when presented with a stimulus of nearly its
own frequency, which would certainly happen in the gre-
garious species, he discovered the jamming avoidance
response. Presented with a sinusoidal stimulus near its own
frequency, the fish either accelerates or decelerates its own
Ž.
Fig. 2. Patterns of electric organ discharges in teleosts. A An electric
catfish, Malapterurus electricus, activity recorded head positivity up-
ward. Mechanical stimulation evoked a train of five pulses with a
ŽX.
maximum frequency of ;190rs. A A single pulse could also be
Ž.
evoked. Recorded at a faster sweep speed. B–D Discharge of weakly
electric gymnotids, South American fishes, recorded head positivity
Ž.
upwards. B A pulse fish, Gymnotus carapo, emits pulses at a basal
frequency of ;35rs. Touching the side of the fish at the time indicated
by the downward step in the lower trace caused an acceleration up to
ŽX.
;65rs. B At a faster sweep speed, the single pulses show three
Ž.
phases, initially head negative. C Sternopygus macrurus, a wave fish,
discharges at ;55rs. The horizontal line indicates the zero potential
Ž.
level. The discharge has little DC component. D Sternarchus
()
Apteronotus albifrons, a high frequency wave fish, emits biphasic
pulses at ;800rs. The horizontal line indicates the zero potential level.
wwxx
Calibrations in volts and milliseconds from Ref. 11 .
discharge to increase the frequency difference and thereby
reduce interference. The central pathways and physiology
of this response and of electroreception in general were
extensively and productively explored by Walter Heilegen-
wx
berg, who was tragically killed in an airplane crash 32 .
Walter had many gifted collaborators.
With the background of gross stimulation of the electric
fish and after more or less exhaustingly reporting the
wx
modes of operation of electric organs 8,11 , Emilio Aljure
and I looked in the spinal cord and then medulla of
mormyrid electric fishes. These species are pulse fishes
and generate very brief discharges -0.5 ms in duration.
Since the generating cells, or electrocytes, emit bi- or
triphasic pulses, very precise synchrony is required to
prevent cancellation of out of phase activity. Although the
electromotor neurons were not visualizable in the spinal
cord, it was not that hard to penetrate neighboring cells
and demonstrate electrotonic coupling directly. In these
species, the electromotor neurons showed quite large diam-
Ž
eter dendrodendritic connections and with uniform stain-
.
ing the cells can appear syncytial or multinucleate, Fig. 3 .
Now here was a preparation that one could, without shame,
ask one’s anatomical colleagues to examine. Yasuko Naka-
jima and George Pappas soon showed that there were close
Ž.wx
membrane appositions between the dendrites Fig. 3 17 .
We would now call these structures gap junctions, al-
though the gap was not resolved in the early pictures. We
did suggest that ultrastructural examination could prove
useful for identifying synapses between dendrites where
electrical measurements were hard to obtain. My col-
league, Dominick Purpura, at that time a mammalian
neurophysiologist, did not approve of this suggestion, but
 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%