L’ E x p

40
Correspondances en pelvi-périnéologie - n° 4, vol. II - octobre/novembre/décembre 2002
Exploration
L’
imagerie du cerveau humain est un
apport technologique prometteur pour
l’étude des différentes fonctions périnéales. La
tomographie par émission de positrons (TEP) et
la tomographie d’émission à photon unique
(TEPU) permettent de visualiser le débit san-
guin cérébral (DSC), reflet indirect de l’activité
neuronale, ou d’étudier les systèmes de neuro-
transmetteurs. L’imagerie par résonance
magnétique fonctionnelle (IRMF) fournit des
informations temporelles sur l’activité céré-
brale en réponse à un stimulus donné.
É
TATDES CONNAISSANCES SUR LE CONTRÔLE
NEUROLOGIQUE CENTRAL DE LA CONTINENCE
ET DE LA MICTION
Des études réalisées chez l’animal, ainsi que
des observations cliniques effectuées chez
l’homme (1-4), ont permis de mieux com-
prendre les mécanismes neurologiques impli-
qués dans le contrôle de la miction. Il a ainsi été
individualisé, dès 1920, un centre mictionnel au
niveau de la protubérance, dont la stimulation
électrique permet d’aboutir à une miction har-
monieuse (2, 5). Holstege et al. ont montré que
la stimulation d’une région médiodorsale de la
protubérance provoque, chez le chat, une dimi-
nution de la pression urétrale, un silence élec-
tromyographique périnéal et une contraction
vésicale (6). Cette région, nommée M-region (5,
6), a des connections avec les neurones de la
colonne intermédio-lateralis de la moelle
sacrée. Ces colonnes contiennent des neurones
parasympathiques destinés à l’innervation
vésicale et des interneurones inhibiteurs des
motoneurones innervant la musculature pel-
vienne (noyau d’Onuf). Par l’intermédiaire de
ces projections, la région M est susceptible
d’induire une miction par stimulation du détru-
sor vésical et par inhibition, dans le même
temps, des motoneurones innervant les
muscles pelviens, ce qui se traduit par la relaxa-
tion du sphincter urétral (7). Parallèlement, il a
été démontré qu’une autre région protubéran-
tielle, L-region, avait des projections sur les
motoneurones sacrés des noyaux d’Onuf (7). Il
a été proposé que la région M soit le site
contrôlant la miction, alors que la région L
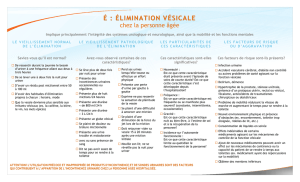
contrôlerait la continence (figure 1).
Malgré leurs rôles respectifs dans le contrôle de
la miction et de la continence, ces deux centres
protubérantiels reçoivent peu de projections
véhiculant l’information sensitive depuis l’ap-
pareil urinaire (8). Cela n’est pas le cas de la
substance grise péri-acqueducale (SGPA) qui
reçoit un grand nombre d’informations sensi-
tives en provenance de la moelle sacrée (9)
(figure 2).Or, il existe des connections entre la
SGPA et les centres mictionnels protubérantiels
(10). Il a ainsi été suggéré que, au cours du rem-
plissage vésical, la SGPA exerçait une influence
tonique permanente sur les neurones du centre
L. Lorsque la vessie atteint un certain seuil de
remplissage et que la situation est appropriée,
il y aurait une permutation de l’activité des neu-
rones du centre L aux neurones du centre M. Il
résulterait de cette modification une relaxation
du sphincter urétral et une contraction du
détrusor, d’où une vidange vésicale complète.
Les études réalisées chez des patients ayant eu
des lésions corticales antérieures soulignent
l’importance du contrôle supra-protubérantiel
de la miction. Les anévrismes artériels céré-
braux antérieurs, les tumeurs et accidents vas-
culaires cérébraux touchant les lobes frontaux
peuvent se compliquer de symptômes urinaires
secondaires à des mictions incontrôlées mais
complètes (11). Bien que leurs rôles restent
* Groupe de recherche de l’appareil digestif,
environnement et nutrition (ADEN),
hôpital Charles-Nicolle, CHU Rouen,
76031 Rouen Cedex.
e-mail : [email protected]
Apport de l’imagerie
médicale à l’étude
de la fonction vésico-
sphinctérienne
■
■
A.M. Leroi*

41
Correspondances en pelvi-périnéologie - n° 4, vol. II - octobre/novembre/décembre 2002
encore mal connus, d’autres structures supra-
protubérantielles telles que le cervelet et l’hy-
pothalamus interviennent également dans le
contrôle neurologique de l’appareil vésico-
sphinctérien.
É
TUDE NON INVASIVE DES MÉCANISMES
NERVEUX IMPLIQUÉS DANS LA MICTION
Plusieurs études ont utilisé l’imagerie fonction-
nelle (TEP ou TEPU) pour comparer les débits
sanguins cérébraux mesurés avant la miction,
vessie vide et pleine, puis pendant la miction,
et enfin après la miction (12-15). Ces études ont
été réalisées chez des sujets volontaires sains
de sexe masculin (12, 13, 15) ou féminin (14),
mais tous droitiers. Pour certains, les régions
cérébrales étudiées s’étendaient de la protubé-
rance au gyrus cingulaire (13, 14). Certaines
zones, telles que le cortex cérébral, n’étaient
donc pas incluses dans leurs études. Enfin,
Nour et al. ont contrôlé par un bilan urodyna-
mique la présence d’une contraction vésicale
au moment de la miction (15).
Il a ainsi été démontré que l’organisation du
système nerveux impliqué dans le réflexe mic-
tionnel était probablement très proche de celui
précédemment décrit chez le chat. Plusieurs
structures du système nerveux central seraient
concernées par la réalisation d’une miction
(figure 3).
Le tegmentum pontique
Comme chez le chat, il a été individualisé deux
zones protubérantielles susceptibles de partici-
per au contrôle nerveux de la miction (12-15). La
première zone, située dans la partie postéro-
médiane de la protubérance, proche du qua-
trième ventricule, serait l’équivalent de la région
M, puisque le débit sanguin cérébral y est signi-
ficativement augmenté lors de la miction (13,
14). Au contraire, la deuxième zone, située dans
la protubérance mais dans une position ventro-
latérale, était activée chez les sujets n’ayant pu
uriner sur ordre (13, 14). Cette zone pourrait cor-
respondre à la région L. En effet, dans les études
de Blok et al. (13, 14), les sujets volontaires qui
ne pouvaient initier de miction, probablement
en raison d’une inhibition psychologique,
contractaient leur sphincter urétral bien plus
fortement que lors du scanner effectué pendant
la retenue, alors que leur vessie était pleine et
qu’ils essayaient d’uriner (figure 3).
Vessie
Aire pré-optique
Substance grise
péri-acqueducale
Afférences
vésicales
secondaires
Aire pré-optique
Substance grise
péri-acqueducale
Centre mictionnel
protubérantiel
L-region
vessie
Sphincte
r
urétral
externe
Noyau d’Onuf
Motoneurones
vésicaux
Interneurone inhibiteur
Figure 1. Schéma
illustrant les
structures spinales
et supra-spinales
impliquées dans les
voies efférentes du
contrôle de la
miction (26). Les
lignes continues
représentent les
projections
excitatrices ; les
lignes discontinues
des interneurones
inhibiteurs sacrés au
noyau d’Onuf
représentent les
voies inhibitrices.
Les voies ne sont
représentées que
d’un côté.
Abrévations : BC,
brachium conjonctivum ;
CA, commissure
antérieure ; IC, colliculus
inférieur ; OC, chiasma
optique ; PON,
protubérance ; SC,
colliculus supérieur ; S2,
second segment sacré.
Figure 2. Schéma
illustrant les
structures spinales
et supra-spinales
impliquées dans les
voies afférentes
véhiculant
l’information sur le
remplissage vésical
(26). Les voies ne
sont représentées
que d’un côté.
Abrévations : BC,
brachium conjonctivum ;
CA, commissure
antérieure ; IC, colliculus
inférieur ; OC, chiasma
optique ; PON,
protubérance ; SC,
colliculus supérieur ; S2,
second segment sacré.

La substance grise péri-acqueducale
(SGPA)
Il a été observé une augmentation significative
du flux sanguin cérébral au niveau de la SGPA
pendant la miction des sujets volontaires sains
de sexe masculin (13). Ce résultat n’a pas été
retrouvé chez la femme (14). Compte tenu du
rôle connu de la SGPA au cours des mictions
chez le chat, ces résultats suggèrent que cette
zone jouerait également un rôle important lors
de la miction chez l’homme. On peut supposer
que l’activation de la SGPA, résultant de la sti-
mulation des tensio-récepteurs vésicaux par le
remplissage vésical, soit responsable de l’acti-
vation du centre M entraînant la miction.
Néanmoins, Nour et al. (15) n’ont pas mis en
évidence d’activation significative de la SGPA
vessie pleine versus vessie vide. Les auteurs en
concluent que soit l’activation de cette région
n’était pas détectable par le TEP, soit la disten-
sion de la vessie dans leur étude était insuffi-
sante pour produire une telle activation. L’autre
hypothèse est que, l’ordre de miction étant
donné, la SGPA serait activée par des voies des-
cendantes provenant de structures supra-pro-
tubérantielles (par exemple, l’hypothalamus),
et activerait à son tour le centre mictionnel pro-
tubérantiel, aboutissant à une miction harmo-
nieuse.
L’hypothalamus
Au vu des résultats de ces études (12-15), l’hy-
pothalamus paraît également être impliqué
dans la miction, ce qui a déjà été démontré chez
le chat (16). Chez cet animal, la stimulation de
certaines structures cérébrales telles que le
gyrus cingulaire antérieur, le noyau pré-optique
de l’hypothalamus ou l’amygdale entraînent
des contractions vésicales (17). Toutes ces
structures donnent naissance à des voies des-
cendantes se projetant sur la SGPA et sur le sys-
tème nerveux dit “émotionnel”. Seul le noyau
pré-optique envoie des projections directement
sur le centre mictionnel protubérantiel
(région M). Bien que la participation du noyau
pré-optique hypothalamique dans la miction
soit confirmée par les études par TEP, son rôle
exact demeure méconnu. On peut supposer
que l’influence directe de cette aire hypothala-
mique sur le centre mictionnel protubérantiel
détermine le début de la miction. Qu’il y ait ou
non miction dépend toujours de l’environne-
ment dans lequel se situe l’individu. Ainsi,
lorsque l’information de plénitude vésicale est
véhiculée via les afférences sensitives d’origine
sacrée et la SGPA et qu’il peut y avoir miction,
celle-ci ne se produit que lorsque certaines
structures, telles que le noyau pré-optique
hypothalamique, ont “décidé” que l’environne-
ment s’y prêtait (13).
Le cervelet
Les études effectuées chez l’homme souffrant
d’ataxie cérébelleuse (18) et les investigations
réalisées chez l’animal après résection du cer-
velet (19) ont montré que le cervelet exerçait
une action inhibitrice lors de la phase de conti-
nence et un rôle facilitateur durant la miction.
Chez le sujet volontaire sain, Nour et al. (15) ont
confirmé l’implication du cervelet dans le
contrôle neurologique de la miction puisqu’ils
ont mis en évidence une augmentation du débit
sanguin cérébelleux au cours de la miction.
Le cortex
Chez l’homme, la miction s’accompagne d’une
augmentation du débit sanguin cérébral dans le
42
Correspondances en pelvi-périnéologie - n° 4, vol. II - octobre/novembre/décembre 2002
Exploration
Figure 3. Partie supérieure : sections horizontales montrant les aires cérébrales activées au cours
de la miction (26). (a) M-region ou centre mictionnel protubérantiel ; (b) substance grise péri-acque-
ducale ; (c) hypothalamus ; (d) gyrus frontal inférieur ; (e) gyrus cingulaire antérieur. L, côté gauche;
R: côté droit du cerveau.
Partie inférieure : sections horizontales montrant les aires cérébrales activées lorsque les volon-
taires essayaient d’uriner sans succès. (a) L-region ou centre protubérantiel de continence ; (b)
gyrus frontal inférieur : L, côté gauche du cerveau ; R, côté droit du cerveau.

cortex sensorimoteur et l’aire motrice supplé-
mentaire du cortex frontal (12, 15). Cependant,
bien que les modifications de la pression abdo-
minale enregistrée lors de l’étude de Nour et al.
soient modestes, la relation entre ces modifica-
tions et l’activité cérébrale objectivée reste pos-
sible. Le foyer d’activité cérébrale déclenchée
par la miction étant très proche des aires acti-
vées lors des efforts de contraction des muscles
abdominaux et des différents muscles du plan-
cher pelvien (20), il n’est en tous les cas pas
possible d’exclure formellement cette hypo-
thèse.
Blok et al. (13) décrivent deux zones corticales
impliquées dans la miction visualisées en TEP.
Dans cette étude, la partie dorso-latérale du
cortex préfrontal droit était activée lorsque la
miction débutait, mais également lorsqu’elle
était permise par le sujet mais non réalisée
(figure 3).Le flux sanguin cérébral était dimi-
nué au niveau du gyrus cingulaire antérieur
droit lorsque le sujet se retenait d’uriner. Cette
désactivation du gyrus cingulaire antérieur
reflèterait une inhibition des informations sen-
sitives vésicales dans le but d’atténuer la sen-
sation de plénitude vésicale et, par conséquent,
le besoin impérieux d’uriner. Il est probable que
l’activation du cortex préfrontal et du gyrus cin-
gulaire antérieur ne soit pas spécifique de la
miction, mais soit liée à des actions plus géné-
rales telles que l’attention et la sélection d’une
réponse appropriée. Ainsi, les lésions du gyrus
cingulaire antérieur sont susceptibles de provo-
quer une incontinence urinaire (11). Cela peut
être dû à un manque d’attention du patient qui
devient incapable de reconnaître la sensation
de plénitude vésicale et n’a donc plus un com-
portement mictionnel approprié.
L’étude de Blok et al. (13) a montré que les
zones cérébrales (cortex et protubérance)
impliquées dans la miction se situaient de
façon prédominante à droite chez des sujets
volontaires sains, tous droitiers. Cette latérali-
sation du contrôle mictionnel permettrait d’ex-
pliquer pourquoi l’incontinence urinaire est
particulièrement fréquente après les lésions
hémisphériques droites (21). Par ailleurs, la
prédominance de l’hémisphère droit dans le
contrôle des fonctions autonomiques a déjà été
démontrée par des études utilisant le TEP et le
TEPU pour le contrôle de la respiration (22) et
de l’orgasme masculin (23). Toutefois, il faut
souligner que les autres études effectuées en
imagerie fonctionnelle sur le contrôle nerveux
de la miction ne confirment pas ce résultat (12,
15).
C
OMPARAISON DIRECTE ENTRE
LA PERCEPTION DU REMPLISSAGE VÉSICAL
ET L
’
ACTIVITÉ NEURONALE
Alors que les précédentes études étaient
essentiellement focalisées sur la miction,
Athwal et al. (24) se sont intéressés à l’activité
cérébrale pour différents volumes de remplis-
sage vésical, non douloureux, ainsi que lors de
besoins mictionnels impérieux. Onze sujets
volontaires sains de sexe masculin (trois gau-
chers et huit droitiers), âgés de 19 à 54 ans, ont
eu un remplissage vésical par cathétérisme uré-
tral. Une TEP était effectuée après chaque rem-
plissage. Le sujet avait ordre de coter sa per-
ception du remplissage vésical (0 = pas de
perception ; 1 = première sensation ; 2 = pre-
mier besoin impérieux ; 3 = besoin impérieux
important ; 4 = besoin impérieux très impor-
tant) avant et après la réalisation du scanner.
Le débit sanguin cérébral de la SGPA augmen-
tait parallèlement au degré de remplissage
vésical, confirmant le rôle, discuté ci-dessus, de
cette structure nerveuse dans le contrôle mic-
tionnel. Une activité protubérantielle était éga-
lement mise en évidence au cours du remplis-
sage vésical. Cependant, contrairement à ce
que l’on aurait pu attendre, cette zone d’hyper-
activité protubérantielle semblait différente de
la région L sollicitée lors des efforts de retenue.
Au cours du remplissage vésical, il a été
observé une activation bilatérale des lobes
latéraux du cervelet et des aires frontales, sug-
gérant que ces différentes structures ont un
rôle non seulement moteur au cours de la mic-
tion (voir ci-dessus), mais également sensitif
pendant le remplissage vésical et le maintien
de la continence. Le débit sanguin cérébral au
niveau du gyrus cingulaire était également
modulé en fonction du remplissage vésical et
de la sensation d’impériosité mictionnelle, et
cela en dehors de toute perception doulou-
reuse. Ce résultat confirme le rôle complexe
joué par le gyrus cingulaire dans l’utilisation
des informations sensitives vésicales afin d’in-
fluencer la perception de besoin impérieux et la
continence urinaire. Aucune modification signi-
ficative d’activité n’a été observée au niveau du
43
Correspondances en pelvi-périnéologie - n° 4, vol. II - octobre/novembre/décembre 2002

44
Correspondances en pelvi-périnéologie - n° 4, vol. II - octobre/novembre/décembre 2002
Exploration
cortex somesthésique lors du remplissage vési-
cal ou de la perception d’un besoin mictionnel
impérieux.
Les structures cérébrales impliquées dans la
perception du besoin impérieux semblent dif-
férentes de celles impliquées dans la sensation
de remplissage vésical. Ainsi, certaines modifi-
cations telles que la désactivation au niveau de
l’hypothalamus, du cortex prémoteur, de cer-
taines parties du gyrus cingulaire de façon bila-
térale étaient corrélées à la perception d’un
besoin mictionnel impérieux et indépendantes
de la perception de plénitude vésicale.
D’autres études (13, 15) ont rapporté l’activa-
tion de l’insula et de l’opercule pendant le rem-
plissage vésical. La stimulation de l’insula chez
l’homme provoque une augmentation du tonus
sympathique (25). L’activation des fibres sym-
pathiques est connue pour accroître la relaxa-
tion du détrusor, entraînant une augmentation
de la capacité vésicale. Ces résultats suggèrent
que l’insula pourrait être une zone d’intégration
de la réponse autonomique au cours du rem-
plissage vésical.
C
ONCLUSION
L’imagerie cérébrale fonctionnelle fournit un
outil unique pour mieux comprendre le contrôle
neurologique de la continence et de la miction
et pour identifier le substrat anatomique de
chacune de ces fonctions. Toutefois, l’attribu-
tion d’une fonction à une région spécifique du
cerveau nécessitera la confrontation des résul-
tats de plusieurs études utilisant des méthodo-
logies variées (sujets volontaires sains et
patients).
RÉFÉRENCES BIBLIOGRAPHIQUES
1. De Groat WC. Nervous control of the urinary
bladder of the cat. Brain Research 1975 ; 87 : 201-
11.
2. Griffiths D, Holstege G, Dalm E, de Wall H.
Control and coordination of bladder and urethral
function in the brainstem of the cat. Neurourol
Urodyn 1990 ; 9 : 63-82.
3. Blok BFM, Holstege G. The central control of
micturition and continence: implications for uro-
logy. BJU Int 1999 ; 83 (Suppl 2) : 1-6.
4. Fowler CJ. Neurological disorders of micturi-
tion and their treatment. Brain 1999 ; 122 : 1213-
31.
5. Barrington RJ. The effect of lesions of the hind
and midbrain on micturition in the cat. Q J Exp
Physiol 1925 ; 15 : 81-102.
6. Holstege G, Kuypers HG, Boer RC. Anatomical
evidence for direct brainstem projections to the
somatic motoneuronal cell groups and autonomic
preganglionic cell groups in cat spinal cord. Brain
Res 1979 ; 171 : 329-33.
7. Holstege G, Griffiths D, de Wall H, Dalm E.
Anatomical and physiological observations on
supraspinal control of bladder and urethral
sphincter muscles in the cat. J Comp Neurol 1986 ;
250 : 449-61.
8. Blok BF, de Weerd H, Holstege G. Ultrastructural
evidence for a paucity of projections from the lum-
bosacral cord to the pontine micturition center or
M-region in the cat: a new concept for the organi-
zation of the micturition reflex with the periaque-
ductal gray as central relay. J Comp Neurol 1995 ;
359 : 300-9.
9. Vanderhost VG, Mouton LJ, Blok BF, Holstege G.
Distinct cell groups in the lumbosacral cord in the
cat project to different areas in the periaqueductal
gray. J Comp Neurol 1996 ; 376 : 361-85.
10. Blok BF, Holstege G. Direct projections from
the periaqueductal gray to the pontine micturition
center (M-region). An anterograde and retrograde
tracing study in the cat. Neurosci Lett 1994 ; 166 :
93-6.
11. Andrew J, Nathan PW. Lesions of the anterior
frontal lobes and disturbances of micturition and
defecation. Brain 1964 ; 87 : 233-62.
12. Fukuyama H, Matsuzaki S, Ouchi Y,
Yamauchi H, Nagahama Y, Kimura J, Shibasaki H.
Neural control of micturition in man examined
with single photon emission computed tomogra-
phy using 99mTc-HMPAO. Neuroreport 1996 ; 7 :
3009-12.
13. Blok BF, Willemsen ATM, Holstege G. A PET
study on brain control of micturition in humans.
Brain 1997b ; 120 : 111-21.
14. Blok BFM, Sturms LM, Holstege G. Brain acti-
vation during micturition in women. Brain
1998b ; 121 : 2033-42.
15. Nour S, Svaer, Kristensen JKI, Paulson OB,
Law I. Cerebral activation during micturition in
normal men. Brain 2000 ; 123 : 781-9.
16. Tang PC, Ruch TC. Localization of brain stem
and diencephalic area controlling the micturition
reflex. J Comp. Neurol 1956 ; 106 : 213-45.
17. Gjone R, Setekleiv J. Excitatory and inhibi-
tory bladder responses to stimulation of the cere-
bral cortex in cat. Acta Physiol Scand 1963 ; 59 :
337-48.
18. Prati A, Cantoni C, Pieri L, Sacchini P, Savino
A, Arnaudi R et al. Urodynamic aspects in cerebel-
lar degenerative disease. Acta Urol Ital 1993 ; 7 :
175-6.
19. Nishizawa O, Ebina K, Sugaya K, Noto H,
Satoh K, Kohama T, Harada T, Tsuchida S. Effect of
cerebellectomy on reflex micturition in the decere-
brate dog as determined by urodynamic evalua-
tion. Urol Int 1989 ; 44 : 152-6.
20. Blok BFM, Sturms LM, Holstege G. A PET
study on cortical and subcortical control of pelvic
floor musculature in women. J Comp Neurol
1997a ; 389 : 535-44.
21. Kuroiva Y, Tohgi H, Ono S, Itoh M. Frequency
and urgency of micturition in hemiplegic
patients : relationship to hemisphere laterality of
lesions. J Neurol 1987 ; 234 : 100-2.
22. Corfield DR, Fink GR, Ramsay SC, Murphy K,
Harty HR, Watson JDG, Adams L, Frackowiak RS,
Guz A. Evidence for limbic system activation
during CO2-stimulated breathing in man. J
Physiol 1995 ; 488 : 77-84.
23. Tiihonen J, Kuikka J, Kupila J, Partanen K,
Vainio P, Airaksinen J et al. Increase in cerebral
blood flow of right prefrontal cortex in man during
orgasm. Neurosci Lett 1994 ; 170 : 241-3.
24. Athwal BS, Berkley KJ, Hussain I, Brennan A,
Craggs M, Sakabikara R, Frackowiak RSJ, Fowler
CJ. Brain responses to changes in bladder volume
and urge to void in healthy men. Brain 2001 ; 124 :
369-77.
25. Oppenheimer SM, Gelb A, Girvin JP,
Hachinski VC. Cardiovascular effects of human
insular cortex stimulation. Neurology 1992 ; 42 :
1727-32.
26. Blok BF, Holstege G. The central nervous sys-
tem control of micturition in cats and humans.
Behav Brain Res 1998a ; 82 : 119-25.
1
/
5
100%