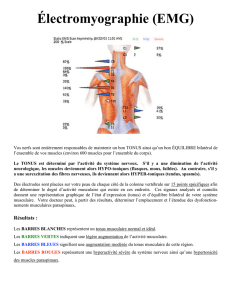
1: van den Tillaar R, Ettema G

1
PHYSIOLOGIE DE L’EQUILIBRE ET DU
DESEQUILIBRE
Le passage de l’environnement marin à la vie terrestre à quatre pattes, puis sur deux jambes, a
nécessité un profond remaniement des systèmes chargés d’assurer l’équilibre. La bipédie est
certainement celui qui a occasionné le plus de changements.
Le sujet assure son équilibre grâce à deux systèmes complémentaires, le premier de type
statique antigravitaire est appelé le tonus postural, le deuxième de type dynamique (maintien de
l’équilibre lors du mouvement) est régulé par l’appareil vestibulaire. Ces deux systèmes adaptent
l’équilibre du sujet en temps réel. Quand le déséquilibre se manifeste la proprioception prend le relais
pour gérer au mieux la situation.
1. TONUS POSTURAL ANTIGRAVITAIRE
Le tonus postural, contrairement au système vestibulaire dont l’action est centrée sur
l’équilibre, a pour objet de maintenir l’organisme érigé grâce aux contractions involontaires des
muscles rachidiens, des ceintures et des membres (ou des membres inférieurs chez les bipèdes). Son
action s’applique donc en priorité sur les muscles extenseurs.
+ Physiologie du tonus postural
Le tonus postural a pour fonction d’assurer le maintien des attitudes de l’animal. Il est assuré
par de nombreuses informations provenant des voies afférentes coordonnées aux différents étages
encéphaliques. Les principales afférences ont pour origine :
= Le réflexe myotatique et le système gamma. Ce réflexe prend naissance
dans les muscles, il est modulé par le système
qui permet la contraction musculaire équilibrante pour
un degré d’étirement optimal. Les afférences musculaires sont identiques aux afférences ostéo-
articulaires, tendineuses et capsulaires. Les informations sont transmises aux centres du tronc cérébral,
des hémisphères et du cervelet. Cet équilibre entre les muscles agonistes et antagonistes permet le
maintien de la posture chez les sujets soumis au champ de la gravitation.
= Le système vestibulaire. Le vestibule (oreille interne) est à l’origine d’un
réflexe tonique dont les centres sont bulbaires cérébelleux et réticulaires. Les voies efférentes
aboutissent aux motoneurone
et
.
= Le système visuel. La vue participe indirectement au tonus postural en
contrôlant la position du corps dans l’espace (placement de la tête, contraction des muscles nucaux).
La perte, même momentanée de la vision au repos, peut entraîner une chute du sujet, si elle n’est pas
compensée par l’un ou l’autre des autres mécanismes assurant le tonus postural.
= Le système ostéo-articulaire cervical est mis en jeu par la variation de la
position de la tête par rapport au tronc. Il s’agit d’un réflexe tonique qui transite par le cervelet et les
centres réticulaires pour aboutir au motoneurone
. Toute contraction anormale des muscles nucaux
(torticolis) peut être à l’origine de sensations vertigineuses.
L’ensemble de ces informations est transmis aux centres nerveux, qui agissent à leur tour sur les
motoneurones
et
. Cinq de ces centres peuvent être individualisés :
Les noyaux gris centraux des hémisphères (néo-striatum et pallidum).

2
Le cortex cérébral, par le biais de la voie cortico-spinale (d’origine prérolandique)
exerce une action activatrice, tandis que la voie cortico-réticulo-spinale agit en
inhibant le système
, et en activant d’autres zones sous corticales.
Le cervelet joue un rôle régulateur sur le système postural en intégrant les
informations cérébro-réticulaires et cérébro-vestibulaires.
L’hypothalamus est inhibiteur dans sa région antérieure, et activateur du système
dans sa région postérieure.
La formation réticulaire du tronc cérébral constitue un véritable carrefour des
informations toniques.
+ Acquisition du tonus postural
Le système postural de nos ancêtres devait présenter des caractéristiques assez semblables à
celles que nous connaissons aujourd’hui chez les vertébrés. Il semble, au vu des études réalisées
auprès d’enfants « sauvages » rapportées depuis plus de 500 ans, que le tonus postural subisse chez
l’homme les effets d’un apprentissage à la bipédie (l’enfant de Hesse marchait à quatre pattes et était
dans l’incapacité de se redresser, de même les enfants de Lituanie, d’Hanovre, des Pyrénées, de
Salzburg, de Sékandra, de Kamala... n’ont jamais réussi à retrouver une bipédie satisfaisante).
Autrement dit, les connexions indispensables au bon fonctionnement du tonus postural, s’adapteraient
en fonction de la demande de l’individu lors de la première année, période d’acquisition de la marche
bipède chez les Homo.
Si l’équilibre et le tonus permettent dès les premières semaines le maintien de la tête, puis la
marche à quatre pattes et la station assise, il faut attendre près de 12 mois pour qu’un début de
déplacement bipède apparaisse. Ce dernier, peut d’ailleurs, s’il n’est pas l’objet de stimulation par les
parents, ne jamais apparaître. Tout se passe comme si l’inné ne concernait que le schéma de base sans
donner le mode d’emploi du système. En l’absence de l’apprentissage des parents, l’enfant se trouve
dans l’incapacité d’adapter son tonus postural à la bipédie. Le système postural à disposition de tout
nouveau-né sera donc modulé en fonction des stimulations données par les parents ou l’entourage.
Sans modèle le système est incapable de fonctionner et de s’adapter. Autrement dit, sans parents
bipèdes, il n’y a pas de bipédie. C’est pourquoi, si Tarzan a pu acquérir assez vite une brachiation de
bonne qualité du fait d’un apprentissage donné par les singes, il est certain que Rémus et Romulus,
élevés par la fameuse Louve romaine, n’auraient jamais pu marcher autrement qu’à quatre pattes.
Dans les deux cas la légende dépasse largement les capacités d’adaptation de l’organisme aux
contraintes de la gravitation.
Chronologie de l’installation du tonus postural :
= Tonus prénatal. La mise en place du tonus postural commence in-utéro. Les
changements de position du fœtus dans le liquide amniotique répondent aux stimulations utérines. Dès
la sixième semaine les muscles axiaux du cou et du tronc peuvent répondre aux contractions utérines.
La maturation du système vestibulaire permet les retournements et un début d’orientation. Ce
phénomène est retrouvé chez tous les mammifères.
= De 0 à 3 mois. La mise en place du système postural se fait de la région
encéphalique vers la région caudale. Après le réflexe oculomoteur, ce sont les muscles nucaux, puis
ceux du tronc et de l’abdomen qui sont l’objet d’une maturation neurologique, pour terminer par ceux
des membres inférieurs. Entre la naissance et trois mois, les fonctions psychomotrices d’ajustement et
d’éveil arrivent à maturité. L’enfant commence dès cette date à intégrer les attitudes posturales de ses
parents.
= De 3 à 4 mois. Pendant cette période, l’enfant commence à contrôler ses muscles
oculomoteurs, sa tête s’équilibre, il est capable de suivre un objet sans loucher. Les connexions
cérébelleuses entre les afférences visuelles et nucales se mettent en place.

3
= De 4 à 6 mois. L’enfant passe de la reptation, parfois chaloupée, à une marche à
quatre pattes bien coordonnée. Il tient la position assise du fait de la bonne tonicité des muscles axiaux
et de la coordination oculomotrice.
= De 5 à 8 mois. Lors de la position assise, l’enfant peut se pencher en avant sans
déséquilibre. Le contrôle des ceintures due à la maturation du système pyramidal lui permet de jouer
avec ses mains et de bouger la tête sans perdre l’équilibre.
= De 9 à 12 mois. A partir de cette période, le tonus postural lui permet de se tenir
debout, appuyé contre un support, mais si l’on occupe son regard (modification de la position de la
tête) il tombe par fléchissement des membres inférieurs. Entre 9 et 12 mois il réalise ses premiers pas,
d’abord aidé, puis seul. Ce simple déplacement nécessite une maturation du système réticulé, des
corps striés, du cervelet, du thalamus et du système pyramidal.
= De 1 à 3 ans. Pendant cette période, l’enfant acquière l’équilibre, coordonne le
déplacement de ses bras et de ses jambes et met en place la course. La mise en jeu de l’ajustement
postural, à partir du schéma corporel inconscient géré par les réflexes proprioceptifs, prend une place
prépondérante à cet âge.
= De 3 à 7 ans. Le schéma corporel, parfaitement inconscient jusqu'à ce stade du
développement, commence progressivement à intégrer un niveau conscient par la mise en jeu de la
fonction d’intériorisation. L’ajustement tonique postural est plus précis grâce aux capacités
d’inhibition corticale qui permet d’éliminer les tensions parasites. C’est à ce niveau que débute
l’hominisation du tonus postural. D’inconsciente, l’ensemble de la chaîne motrice se trouve
maintenant soumise à un contrôle cortical, il est possible d’en modifier significativement les réponses.
L’apprentissage devient possible (natation, saut...) *.
L’acquisition du tonus postural tel que nous le connaissons chez Sapiens sapiens est donc
passé par des phases d’apprentissage transmises de génération en génération depuis l’apparition de la
bipédie. Les ajustements permettant le passage d’une position semi-érigée à une position érigée n’ont
certainement pas posé de grandes difficultés d’adaptation posturale du fait de l’existence au sein de
notre système nerveux des structures propres à prendre en compte ces modifications. La question qui
se pose est de savoir si le passage de la position quadrupède à une démarche bipède (semi-érigée, ou
totalement érigée) a nécessité des aménagements plus complexes. L’étude du tonus postural des
primates quadrupèdes ne montre pas sur le plan macro anatomique de véritable spécificité. C’est plus
au plan des connexions neuronales, des localisations ostéo-articulaires des récepteurs et des muscles
effecteurs que se trouvent les différences essentielles. Mais là encore il s’agit vraisemblablement de
processus adaptatifs du même type que ceux mis en jeu au niveau des articulations du membre
inférieur.
En conclusion le tonus postural, comme la bipédie, ne constitue pas un caractère probant
d’hominisation. Seule la maîtrise volontaire du tonus posturale, telle qu’elle peut être acquise après
l’âge de sept ans, peut différencier l’homme des autres primates.
* Ce mécanisme peut être perturbé chez l’adulte dès l’instant où la nuque se trouve heurtée brutalement par un choc direct ou
indirect. Lors de la pratique de la boxe anglaise un coup violent porté à la face peut rompre la chaîne du tonus postural en
provoquant une déconnexion momentanée du système nucal. La chute du sportif est tout à fait significative de cette
interruption tonique. Les genoux fléchissent donnant l’impression que le sportif tombe assis. Dans ce type de KO la
conscience ne se trouve à aucun moment perturbée.

4
2. L’EQUILIBRE
+ L’appareil vestibulaire
L’appareil vestibulaire est apparu chez les vertébrés dès la sortie de l’eau, prenant ainsi le
relais du système latéral qui caractérise tous les vertébrés inférieurs aquatiques (poissons, certains
amphibiens...). Le système latéral présente de multiples fonctions (perception, équilibration...) Il
disparaît chez la presque totalité des amphibiens, mais peut persister de façon plus ou moins complète
en fonction du mode de vie de l’animal (totalement aquatique, aquatique pour la ponte...). Cet organe a
totalement régressé chez les vertébrés supérieurs.
Le labyrinthe, situé au niveau de l’oreille interne, est retrouvé chez tous les vertébrés depuis
les Poissons et les Cyclostomes. Chez les tétrapodes, ce système uniquement relié à l’équilibre de
l’animal se trouve couplé au système auditif par adjonction d’une cavité supplémentaire, l’oreille
moyenne.
Le labyrinthe membraneux est issu de cellules épiblastiques qui s’invaginent pour donner
l’otocyste.
C’est à partir des parois de cette vésicule que naissent les neurones ganglionnaires à l’origine
de la mise en place du nerf stato-acoustique. L’otocyste se divise en deux parties qui donneront dans la
moitié dorsale la cavité de l’utricule et dans la partie ventrale le saccule. L’utricule donne naissance
aux trois canaux semi-circulaires qui s’orientent dans l’espace suivant les trois plans d’un trièdre
rectangle. L’extrémité de chacun de ces canaux se renfle pour donner l’ampoule.
Le saccule s’évagine pour former la lagéna qui donne naissance au canal endolymphatique.
Parallèlement à ces transformations morphologiques l’épithélium du labyrinthe se différencie en
plages sensorielles (cellules ciliées) dont les fonctions varient suivant leur localisation. On retrouve :
. Une papille basilaire acoustique uniquement présente chez les tétrapodes
. Des macules statiques (utriculaire, sacculaire et lagénaire) chargées de sécrétions calcaires
(otoconies ou otolithes).
. Des crêtes moins mobiles, dépourvues d’otoconies placées dans les ampoules des canaux
semi-circulaires.
= L’oreille interne humaine
Chez l’homme, l’appareil vestibulaire se trouve localisé à l’intérieur du rocher. Les canaux
semi-circulaires sont orientés à 30° vers l’arrière pour le canal horizontal (ou canal externe), à 45° par
rapport an plan sagittal pour le canal vertical antérieur, et à 90° par rapport au précédent pour le canal
vertical postérieur. Chacun de ces canaux communique avec l’utricule qui s’ouvre lui-même dans le
saccule. Ces cavités contiennent de l’endolymphe dont les déplacements sont en rapport direct avec les
mouvements de la tête. Les accélérations et décélérations stimulent spécifiquement les crêtes
ampullaires localisées dans les canaux semi-circulaires, les changements de posture stimulent
électivement les macules. Ces récepteurs donnent naissance à des fibres convergeant vers le conduit
auditif interne où sont localisés les premiers neurones (ganglion de Scarpa). Les prolongements de ces
cellules donnent naissance au nerf vestibulaire qui s’associe au nerf cochléaire pour donner la VIIIème
paire crânienne. A l’intérieur du tronc cérébral ces fibres se connectent avec celles du deuxième
neurone (noyaux vestibulaires). Les fibres issues des noyaux vestibulaires se dirigent vers le cervelet,
la corne antérieure de la moelle, les noyaux oculomoteurs (paires III, IV et VI).
L’équilibre des vertébrés terrestres a, depuis la sortie des eaux, eu pour références la verticale,
direction d’application des forces gravitaires, et l’horizontale point de repère du système visuel. Si
l’oreille moyenne a subit une évolution structurale considérable, avec la migration et la mise en place
des osselets, l’oreille interne ne s’est que faiblement transformée. Néanmoins, l’étude des pièces
osseuses protégeant le système vestibulaire a permis d’évaluer chez les fossiles la taille de cet organe,
l’orientation des canaux semi-circulaires et la position relative de ces canaux les uns par rapport aux

5
autres. Une étude récente portant sur ce sujet montre que l’augmentation globale du système
vestibulaire, et notamment la grandeur du rayon de courbure du canal postérieur, était proportionnelle
à la taille du crâne considéré. La seule différence notable porte sur les néandertaliens qui présentaient
un canal postérieur placé plus bas que celui des Homo (habilis, erectus et les autres…). Cette
disposition particulière peut être intéressante pour caractériser ce cousin et évaluer son éloignement de
la branche sapienne, mais ne joue sans doute aucun rôle physiologique particulier. Les néandertaliens
répondaient de façon identique aux contraintes de l’équilibre.
La modification de l’équilibre au cours de notre courte évolution, au regard des temps
préhistoriques, se situe donc au moment de l’acquisition de la bipédie qui se trouve être par définition
une position instable, non gérable en totalité par le tonus postural. L’étude de cette position instable
montre que le déséquilibre est essentiellement antéropostérieure. Le pendule inversé (point d’appui au
niveau des chevilles) oscille donc d’avant en arrière et maintient son équilibre grâce à la contraction
des muscles agonistes/antagonistes de la jambe suivant un schéma de contraction partant du bas et se
déplaçant vers le bassin. La perte de cette fonction (perte de la proprioceptivité, vieillissement,
immobilisation prolongée...) tend à déplacer le point fixe pendulaire vers le bassin, rendant l’ensemble
particulièrement instable. Cette observation s’articule parfaitement avec la genèse de la bipédie,
acquise suivant un plan antéro-postérieur tandis que les membres postérieurs gardaient latéralement
une fonction pratiquement identique à celle de la quadrupédie. L’acquisition de l’équilibre par
l’enfant, après celle de son tonus posturale, répond certainement à un apprentissage mené de front avec
ce dernier dès les premiers jours de la vie. Contrairement au tonus qui commence à se développer de
façon réflexe in utero, l’apesanteur relative à laquelle est soumis l’enfant pendant la grossesse ne
permet sans doute pas une mise en place aussi rapide des connexions neuronales spécifiques à la
fonction équilibratrice. La maîtrise de l’équilibre sur les pieds, puis sur un pied, les yeux ouverts puis
fermés, éventuellement sur les mains pour certains sportifs, s’acquière par l’entraînement, c’est à dire
la coordination des réponses motrices à partir des stimuli efférents. Le ski, le skate bord, la planche à
voile... sont autant de situations de déséquilibres d’autant plus facilement à gérer que l’on est jeune.
La boxe, déjà citée dans le cadre du tonus postural donne encore ici un exemple caricatural de
l’atteinte de l’équilibre par un traumatisme. Si le coup porté est un crochet, les canaux semi-circulaires
de l’oreille interne subissent une accélération rotatoire rapide qui perturbe l’état d’équilibre du boxeur.
Le sportif chute, et se trouve dans l’incapacité de se relever du fait de la perte d’équilibre (on le voit
tituber, retomber du fait d’un vertige qui se calmera spontanément en quelques minutes, bien souvent
après avoir été compté dix). L’équilibre n’apparaît donc pas comme un facteur sélectif propre à
infléchir les processus évolutifs, mais comme une fonction modulable que les premiers bipèdes
utilisèrent à leur profit pour modifier leur système de locomotion.
3. PROPRIOCEPTION
La proprioception peut se définir comme l’ensemble des moyens neuromusculaires
inconscients permettant de rétablir dans un temps très rapide une situation de déséquilibre. La
proprioception concerne l’ensemble des réactions d’équilibre de l’organisme ainsi que le tonus
postural. Elle associe l’oreille interne et les yeux, mais également l’ensemble des informations
neuromusculaires susceptibles de donner une information instantanée sur la position de notre corps
dans l’espace. Elle correspond physiologiquement aux voies profondes de la sensibilité. La
proprioception est donc directement en relation avec la vue, la position de la tête dans l’espace et le
système neuromusculaire. Les organes sensoriels mettant en jeu ce système sont placés aux niveaux du
crâne (œil et oreille interne), de la nuque et de l’ensemble des récepteurs proprioceptifs articulaires,
ligamentaires, musculaires et cutanés. Les relais nerveux sont réalisés au niveau du tronc cérébral, du
cortex et surtout du cervelet. Les informations efférentes sont d’ordre moteur et concernent
essentiellement les muscles de la nuque et des membres inférieurs.
Chez l’homme, la proprioception est essentiellement en rapport avec les membres inférieurs et
les muscles chargés du tonus postural. Contrairement à ce dernier, qui est uniquement chargé de
 6
7
8
9
6
7
8
9
1
/
9
100%