Corrigé CB 1 - Poly

1
Corrigé CB 1 (Prépa Orthoptiste)
Question 1 Sujet rédactionnel ( /8)
Le VIH (virus de l’immunodéficience humaine) est un virus qui a pour hôte l’organisme humain.
Comme tout virus, il ne peut réaliser son cycle de développement qu’à l’intérieur des cellules. Le VIH est un
parasite intracellulaire obligatoire responsable du SIDA. Le VIH est un virus qui a pour cellule cible les cellules
du système immunitaire pourvues du marqueur CD4, essentiellement les lymphocytes T4 (LT4) . Il affecte donc
la population de ces LT4.
Comment évolue le taux des lymphocytes T4 de la déclaration de la maladie à la mort du sidéen ?
Quel est le rôle essentiel des lymphocytes T4 dans les défenses de l’organisme ?
Dans un premier temps, nous présenterons les trois phases de l’infection par le VIH et parallèlement nous
étudierons l’évolution du taux de lymphocytes T4 au cours de ces trois phases. Puis dans un second temps, nous
étudierons les mécanismes d’action des LT4 au cours des réactions immunitaires acquises.
I] Les trois phases de l’infection par le VIH et la population de lymphocytes T4.
On reconnaît trois phases dans l’infection par le VIH :
A- La primo-infection.
Sa durée de quelques semaines est marquée par des symptômes souvent discrets et non spécifiques
(fièvre, maux de tête…), qui disparaissent par le suite. Sur le plan biologique, elle est marquée par une
augmentation importante de la charge virale plasmatique, associée à une diminution de la population de LT4.
Mais la charge virale chute rapidement, en même temps que le nombre de LT4 redevient pratiquement normal.
Cela est dû aux réactions immunitaires acquises vis-à-vis du VIH, réactions marquées par l’apparition
d’anticorps plus ou moins neutralisants et de LT8 cytotoxiques. Durant cette phase, le système immunitaire est
donc fonctionnel.
La seconde phase est la phase asymptomatique.
B- La phase asymptomatique.
Elle dure en moyenne une dizaine d’années ; elle est marquée par l’absence de symptômes pathologiques,
ce qui traduit un système immunitaire efficace. En même temps, la charge virale reste faible. Cela masque
cependant la poursuite de l’infection par le VIH, qui se traduit notamment par une lente diminution de la
population de LT4.
La troisième phase est la phase de SIDA déclaré.
C- La phase de SIDA déclaré.
Elle est marquée sur le plan clinique par de multiples infections opportunistes ( sarcome de Kaposi,
pneumonie…) dues à des agents pathogènes (bactéries, virus…) auxquels l’organisme résistait durant la seconde
phase.
Cela traduit une immunodéficience générale, c'est-à-dire l’incapacité d’avoir des réactions immunitaires acquises
et spécifiques vis-à-vis des agents infectieux. Sur le plan biologique, cette phase est marquée par une
augmentation de la charge virale et par un nombre de LT4 inférieur à 200/mm3 de sang.
La défaillance du système immunitaire dans la troisième phase apparaît donc lorsque la population de LT4 est
réduite au cinquième de sa valeur physiologique moyenne (1000/mm3). Dans les deux premières phases, le
nombre de LT4 reste relativement élevé et il n’y a pas d’immunodéficience. De plus, les LB et les LT8 ne sont
pas les cibles du virus et restent présents et fonctionnels. Ainsi, la corrélation entre le nombre très réduit de LT4
et l’immunodéficience traduit l’importance de ces lymphocytes pour la réalisation des réactions immunitaires.
Quels sont les mécanismes d’action des LT4 ?
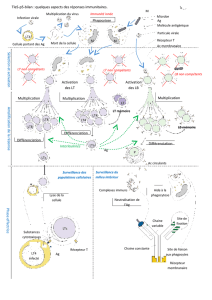
II] Mécanismes d’action des LT4 au cours des réactions immunitaires acquises.
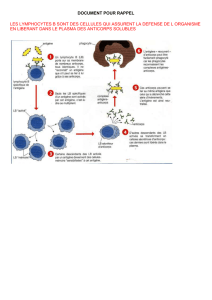
Les cellules effectrices de l’immunité acquise sont les plasmocytes, cellules sécrétrices d’anticorps
plasmatiques spécifiques d’un antigène microbien précis et particulièrement efficaces contres les bactéries
extracellulaires. Les autres cellules effectrices sont les lymphocytes T cytotoxiques qui détruisent spécifiquement
les cellules infectées par un virus (entre autres).
Ces cellules effectrices sont formées à partir de cellules précurseurs, les LB pour les plasmocytes et les LT8 pour
les lymphocytes T cytotoxiques.
A- LB et LT4.
Chaque LB porte des récepteurs, des anticorps membranaires spécifiques d’un antigène donné.
La première étape d’une réaction immunitaire impliquant les LB est la reconnaissance d’un antigène par les
anticorps membranaires spécifiques d’un clone de LB, mais cette reconnaissance préalable indispensable ne
suffit pas pour activer les LB sélectionnés (ou alors cette activation reste très faible), il faut un deuxième signal
d’activation fourni par les LT4. En réponse aux interleukines (messagers chimiques) sécrétées par ces LT4, les

2
LB sélectionnés par l’antigène subissent une expansion clonale (nombreuses multiplications par mitose), et les
cellules qui en sont issues se différencient en plasmocytes et en LB mémoire.
En réalité, ce sont les LT4 spécifiques des antigènes de l’agent pathogène en cause qui sont activés et qui
sécrètent des interleukines ; seuls les LB ayant reconnu ces antigènes expriment des récepteurs aux interleukines,
et peuvent donc subir l’expansion clonale et la différenciation en cellules effectrices.
Les anticorps sécrétés par les plasmocytes peuvent donc se fixer sur le VIH ( dans le cadre de notre sujet) et le
neutraliser en l’empêchant de pénétrer dans une cellule.
B- LT8 et LT4.
Pour les LT8, le principe est le même. Les interleukines sécrétées par les LT4 sont aussi nécessaires à
l’expansion clonale de LT8 ayant reconnu au préalable, par leurs récepteurs membranaires spécifiques
(récepteurs T), la présence dans l’organisme de cellules infectées par le VIH (dans le cadre de notre sujet). Par la
suite, ces LT8 se différencient en LT cytotoxiques.
Les lymphocytes T cytotoxiques entrent en contact avec les cellules infectées par le VIH (dans le cadre de notre
sujet) et entraîne la mort de cette cellule par l’intermédiaire de perforines (protéines capables de créer des pores).
La diminution considérable du nombre de lymphocytes T4 en phase de SIDA déclaré est donc à l’origine de
l’absence de stimulation des clones de LB et LT8 spécifiques des antigènes des agents pathogènes à l’origine des
maladies opportunistes.
Conclusion :
L’immunodéficience de la phase de SIDA déclaré est due à la chute considérable du nombre de LT4, cellules
immunitaires indispensables à la réalisation de presque toutes les réactions de l’immunité acquise. Les LT4 sont
considérés comme les pivots des réactions immunitaires acquises.
Actuellement pour prolonger la vie du patient, on voudrait injecter des lymphocytes T4 de telle façon à reculer la
déclaration de la phase SIDA.
Introduction : 1,5 point.
Conclusion : 1 point.
Transition : 0,5 point.
Plan : 1 point.
Réponse argumentée, orthographe, syntaxe et répondre à la problématique posée : 4 points.
Question 2 Sujet Pondichéry 2010 ( /4)
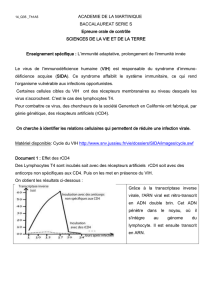
Le but de cet exercice est de confirmer ou infirmer une hypothèse : existe-t-il une relation entre l’évolution
précoce de la charge virale et la survenue d’un SIDA déclaré en l’absence de traitement ? (1 point)
Le document proposé est un histogramme issu d’une étude statistique réalisée à deux stades de l’infection. (0,25
point)
Un an après l’infection, la mesure du degré d’infection encore appelée charge virale a été effectuée. On mesure
la charge virale car le VIH étant un rétrovirus, son information génétique se trouve sous forme d’ARN. Cette
quantité d’ARN représente donc la quantité de virus présents. Dans cette étude, cette quantité virale varie de
1000 à 100000 copies d’ARN/mL (0,5 point)
Cinq ans après l’infection, une étude a été réalisée pour savoir si le patient non traité était entré en phase SIDA
ou non. (0,25 point)
L’étude en parallèle de ces deux données (sur un nombre de patients non précisé) montre une relation entre la
charge virale et l’entrée en phase SIDA déclaré c'est-à-dire l’entrée dans la maladie. (0,5 point)
On constate que si la charge virale se situe entre 1000 à 5000, environ 8% des individus entreront en phase SIDA
déclaré 4 ans plus tard. (0,5 point)
Par contre si la charge virale se situe entre 40000 et 100000, environ 62% des individus entreront en phase SIDA
déclaré quatre ans plus tard. (0,5 point)
Cette étude statistique montre clairement une relation entre la charge virale et le risque de développer le SIDA.
On peut donc confirmer cette hypothèse. (0,5 point)

3
Question 3 « L’atrophie de l’appareil génital chez des souris transgéniques » ( /4)
Les données fournies vont permettre d’expliquer l’atrophie de l’appareil génital chez des souris transgéniques.
Chez les souris mâles transgéniques, l’appareil génital est atrophié. Or, au cours du développement
embryonnaire, la différenciation des voies génitales mâles se fait sous l’action de la testostérone sécrétée par les
cellules interstitielles du testicule fœtal. Le texte indique que l’AMH agit sur les cellules interstitielles. Quelles
sont les modalités de son action ? (1 point)
L’énoncé nous indique que la présence d’AMH entraîne la disparition des canaux de Müller (0,25 point).
Comme les souris transgéniques ont de l’AMH en abondance, on peut supposer que les canaux de Müller sont
atrophiés (0,25 point). D’après nos connaissances, l’autre future voie génitale, le canal de Wolff ne persiste que
si la testostérone est présente (0,25 point). D’après les données écrites du seul document proposé, la synthèse de
testostérone se fait à partir du cholestérol. Cette synthèse nécessite le transporteur StAR et les enzymes E1, E2 et
E3 (0,5 point)
D’après le graphique qui nous est proposé, les molécules sont présentes mais avec des concentrations très
différentes entre elles (0,125 point) . StAR, E1 et E2 sont présentes avec des taux qui varient entre 60 et 90% par
rapport à des souris mâles témoins (0,125 point) . En revanche, E3 n’est présente qu’au taux de 20%.On peut
considérer que le faible taux de E3 limite la synthèse de testostérone ce qui ne permettra pas le maintien des
canaux de Wolff (0,5 point)
On peut expliquer l’atrophie de l’appareil génital chez les souris transgéniques de la manière suivante : la
présence d’AMH, sécrétée par les cellules de Sertoli du testicule, entraîne la disparition des futures voies
génitales femelles, les canaux de Müller. Les cellules interstitielles du testicule devraient synthétiser de la
testostérone, ce qui permettrait le maintien des futures voies génitales mâles, les canaux de Wolff. La très faible
quantité de l’enzyme E3 limite fortement la synthèse de testostérone ce qui empêche le maintien des canaux de
Wolff. (0,75 point)
Ainsi la dégénérescence des canaux de Müller et le non maintien des canaux de Wolff entraînent l’atrophie des
voies génitales. (0,25 point)
Question 4 « Anomalies de la méiose » ( /4)
La méiose permet le passage de la phase diploïde à la phase haploïde. Elle est composée de deux divisions
cellulaires successives qui conduisent à la présence d’un lot haploïde de chromosomes par cellules filles. La
méiose est à l’origine de la formation des gamètes.
Dans certains cas se produisent certaines anomalies au cours de la méiose. (0,5 point)
Nous allons rechercher, à partir de l’analyse du document fourni, où se trouve l’anomalie à l’origine des
caractéristiques des gamètes. (0,25 point)
Le document nous présente trois méioses dont une méiose normale (A). Celle-ci est constituée d’une première
division où a lieu la séparation des chromosomes homologues. Puis une seconde division a lieu, au cours de
laquelle on assiste à une séparation des chromatides. (0,5 point)
Lorsqu’on analyse les gamètes issus de la méiose B, on constate que deux gamètes B1 et B2 ont trois
chromosomes dont deux homologues et les deux autres, B3 et B4, un seul chromosome. (0,25 point) Or, puisque
la cellule qui subit la méiose a quatre chromosomes, les gamètes devraient posséder deux chromosomes comme
ceux issus de la méiose normale. Donc les gamètes issus de la méiose B ont soit un chromosome en trop, soit un
chromosome en moins. (0,5 point)
Comment peut-on expliquer l’anomalie de la méiose B ?
En anaphase de première division de méiose, les deux chromosomes homologues d’une paire vont vers le même
pôle au lieu d’aller l’un vers un pôle de la cellule et l’autre vers l’autre pôle. (0,25 point)Le comportement de
l’autre paire de chromosomes est normal (0,125 point). Il en résulte que les deux cellules issues de cette
première division ont l’une 1 chromosome dédoublé et l’autre trois chromosomes dédoublés. (0,25 point) La
seconde division, caractérisée par la séparation des chromatides de chaque chromosome, est normale d’où
l’obtention de quatre cellules, deux à un seul chromosome et deux à trois chromosomes. (0,25 point)
Lorsqu’on analyse les gamètes issus de la méiose C, on constate que deux des gamètes sont normaux (C3 et C4)
à deux chromosomes, les deux autres sont anormaux, l’un à trois chromosomes, l’autre à un seul chromosome.
(0,25 point)
La première division de la méiose est normale. En revanche, la seconde division d’une des cellules issues de la
première division est anormale. Au cours de cette division, les deux chromatides issues d’un des deux
chromosomes dédoublés vont vers le même pôle de la cellule au lieu d’aller l’une vers un pôle et l’autre vers
l’autre pôle (0,25 point). Le comportement des chromatides issues de l’autre chromosome est normal. (0,125
point) Il en résulte qu’une cellule issue de cette division a trois chromosomes et l’autre un. (0,25 point)
Cet exercice montre que des anomalies peuvent survenir lors de la méiose, lors de la formation des gamètes. Ces
anomalies peuvent être à l’origine de maladies génétiques comme la trisomie 21. (0,25 point)
1
/
3
100%