Recueil de questions d`examen

Recueil de questions d'examen
Cours : Zoologie 2 (Prof. M.Milinkovitch)
U.L.B.
Réponses basées sur :
Cours théorique de Zoo2 (Prof. M.Milinkovitch) ULB
Cours théorique de Biologie Animale BGU04 (?ref)
DEUG en Biologie Animale SVI04 (?ref)
Corrigés des années précédentes
Notes de démonstration de Zoo2 ULB
Thomas Lecocq - 2004
NB : T = Terminé et C = Peut nécessiter un complément

Accrasiales et leur mouvement
Animaux pluri ou unicellulaires ?
Les accrasiales sont des cellules amoeboïdes qui vivent individuellement dans les sols forestiers
et se divisent toutes les trois à quatre heures, se nourrissant de bactéries.
Quand les ressources sont épuisées, l'accrasiale émet de l'cAMP de façon pulsatile.
D'autres accrasiales recoivent ce signal (récepteur membranaire) et se dirigent vers la source en
émettant elles aussi du cAMP. On assiste à une émission concentrique de cAMP. Les accrasiales
se dirigent donc dans une direction, par chémotactitisme et remontent le gradient de
concentration.
De plus, les gènes de synthèse de molécules d'adhésion sont activés. Les accrasiales forment
une limace continue (slime mold) et se regroupent au centre d'aggrégation. Elles forment une
structure pluricellulaire. La structure se dresse en remontant un gradient d'humidité et de
luminosité. Forme une structure élevée, les cellules se recouvrent de cellulose et meurent, sauf
les spores qui pourront être dispersées et germer si les conditions deviennent favorables.
Ils sont donc uni ET pluricellulaires.
* Comment un système où une partie des cellules se sacrifie pour les autres peut-il être viable ?
Seules les accrasiales ayant un patrimoine génétique très proche ressentent l'impulsion d'cAMP.
Ce n'est donc pas contraire à l'évolution que de se sacrifier pour un code générique pratiquement
semblable au sien (a fortiori si ce sont des clones).
T
C
Page 2 sur 75

Adaptation à la nage/saut/vol/reptation
* Adaptation au vol.
On trouve cette adaptation chez des reptiles fossiles (ère secondaire).
a\ Chez les oiseaux.
Pour former l aile, on a un rééquilibrage en longueur des trois segments. Le cubitus est le plus
développé par rapport au radius. On a un allongement de l autopode, l humérus diminue.
Cette adaptation se fait aussi par une disparition des doigts. Le 1 est visible par ses vestiges, les
2 et 3 sont bien développés (ils portent les plumes).
Le membre postérieur s adapte aussi à cette locomotion. Pour supporter le choc de l atterrissage,
le fémur devient horizontal, la cheville se rigidifie par des soudures, l articulation passe au milieu
de l autopode et donne un os unique qui se soude sur le tibia : le tibiatarse. De plus, le métapode
se soude en un os avec le reste du tarse : le tarsométatarse. Seuls les doigts restent libres.
b\ Chez les mammifères (les chiroptères).
Les doigts deviennent prépondérants (seul le doigt 1 se réduit : il permettra de s accrocher au
sol). Ces doigts vont soutendre une membrane (expansion tégumentaire) : le patagium.
* Adaptation à la nage.
On trouve cette adaptation chez les mammifères cétacés. Ils ont une dynamique caudale qui
assure la propulsion. Cette dynamique entraîne une régression de la musculature latérale : il ne
reste qu une musculature dorso-ventrale en relation avec la nageoire caudale horizontale.
Les membres postérieurs disparaissent.
Les membres antérieurs donnent des palettes natatoires. Tous les os de ces membres
s élargissent et se raccourcissent (dans la longueur) : l humérus est court et large, le cubitus et le
radius sont courts et longs.
On a une simplification du basipode. Les cinq doigts sont hyper-phalangés (jusqu à 21
phalanges). Toutes les articulations sont bloquées et des cartilages s accumulent dans ces
articulations sauf dans la basale.
* Adaptation au saut.
Dans ce mode de locomotion, le membre est un ressort à compression. Les trois segments ont
une longueur équivalente (allongement de l autopode). Le système de saut est le même chez les
grenouilles, les kangourous, les lièvres
On a toutefois deux modalités de saut (de réception) :
- Chez la grenouille, sauteur quadrupède, la propulsion est réalisée par les membres postérieurs
et la réception est assurée par les membres antérieurs.
- Chez les kangourous, sauteurs bipèdes, propulsion et réception sont assurées par les membres
postérieurs.
* La reptation.
On a une disparition totale des membres.
T
C
Page 3 sur 75

Adaptation des membres à la course
1\ Le membre plantigrade (le plus primitif).
La marche se fait sur la plante des pieds, tout l autopode repose sur le sol (exemple : l Homme).
L autopode est court et le métapode est bien développé. Cette adaptation permet la marche.
Celle-ci se perfectionne chez l Homme grâce à la bipédie et sert à l assouplissement de la
démarche. On assiste à la création d une voûte plantaire sous chaque pied. Le pied repose sur
trois zones : le talon, le pouce (gros orteil) et le petit orteil.
2\ Le membre digitigrade.
L accélération de la locomotion (trot ou course) se fait par un allongement des membres (plus
grandes enjambées) et par une diminution de la surface au sol. Cette diminution se fait par le
soulèvement de l autopode.
Pour cette digitigradie, le basipode et le métapode se redressent ; le talon est haut, au-dessus du
sol. Seuls les doigts (acropode) touchent le sol.
3\ Le membre onguligrade.
Dans ce type de membre, on parle de digitigradie onguligrade ou d onguligradie.
On a un redressement de l acropode. Tout l autopode se soulève. Seule l extrémité des doigts
touche le sol par la phalangette. C est la caractéristique des ongulés. De plus, on a la formation
de sabots.
Le doigt 1 disparaît toujours et on a au maximum quatre doigts par patte. On a deux lignées
distinctes.
a\ Les mésaxoniens.
L axe du membre passe par le doigt 3 (ordre des périssodactyles : zèbres, chevaux, tapirs). Ce
doigt 3 est toujours plus développé que les autres.
Les tapirs ont quatre doigts à l avant et trois doigts à l arrière (le doigt 5 disparaît).
Les chevaux n ont plus que le doigt 3 avec un sabot. Parallèlement à la disparition des doigts, les
pièces osseuses disparaissent : on a une simplification de l autopode. Il reste des vestiges des
métapodes 2 et 4 sous forme de stylets.
b\ Les paraxoniens.
L axe du membre passe entre les doigts 3 et 4.
Chez l hippopotame, les quatre doigts touchent le sol.
Chez les autres espèces, on a une réduction des doigts externes (2 et 5) :
- Le porc a ses doigts 2 et 5 réduits de telle sorte qu ils ne touchent plus le sol mais forment
deux ergots hauts sur la patte.
- Les cervidés : les doigts 2 et 5 forment deux stylets latéraux.
- Les b ufs et moutons : les doigts 2 et 5 ont disparus ainsi que les structures qui les précèdent
sur le métapode et sur le basipode.
On a une fusion progressive des métapodes 3 et 4 (è l os canon) : c est une caractéristique des
paraxoniens (les arctiodactyles).
T
C
Page 4 sur 75

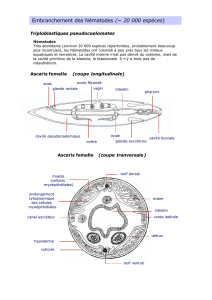
Amphioxus
L'amphioxus est un céphalocordé. Il est benthique et reste fixé dans le sable où il filtre la
nourriture et respire. L'eau entre par la bouche et au niveau des branchies les particules
alimentaires sont attrapées dans un mucus, elles continuent leur trajet au niveau du tube digestif.
L'eau est quant à elle expulsée par les fentes branchiales.
Coupe Transversale :
En CT, l'amphioxus ressemble très fort au chrodé type.
En effet :
- Tube neural dorsal creux,
- chorde
- fentes branchiales
- une queue postnatale
Il possède aussi des myomères de muscles segmentés insérés de manière oblique sur la chorde
qui lui permettent de bouger par ondulations, la corde servant de relais mécanique.
Il y a trois types de cils :
° Dans le vestibule, entouré par les tentacules empéchant les grosses particules de pénétrer dans
le tube digestif il y a des cils qui provoquent un effet de rotation. NB : le vestibule est la partie
antérieure à la bouche entourée par les tentacules. La bouche est une ouverture circulaire dans le
vélum, le bout du vestibule.
° Sur les branchies, ils permettent de créer un courant d'eau et donc de la faire passer sur les
branchies pour réoxygéner le sang. L'eau est ensuite éjectée par les fentes branchiales.
° Sur les sillons ventral (endostyle) et dorsal du pharynx. La nourriture éventuellement contenue
dans l'eau est capturée par un mucus et des cils emmenant le mucus et les aliments vers les
intestins.
T
C
Page 5 sur 75
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
1
/
75
100%