les vegetaux ligneux - CCAVV

Sortie botanique du 16 février 2013
Site Natura 2000 de la reculée des Planches-Près-Arbois
Pascal Collin 2013 page 1
La dichotomie, de gauche à droite : chez le palmier doum (Hyphaene thebaica) – Chez Psilotum, un genre de fougère – Chez une
algue (Fucus vesiculosus).
LES VEGETAUX LIGNEUX
CROISSANCE ET ARCHITECTURE
LA RAMIFICATION
La ramification détermine le port d'un individu. On rencontre deux modalités :
La dichotomie :
Elle est très répandue chez les cryptogames1. Chez les Ptéridophytes2, on notera les
sélaginelles, les lycopodes et surtout Psilotum (image du milieu ci-dessous). La
dichotomie est très rare chez les phanérogames3 (Hyphaene4, Chamaedorea
elegans5
).
La ramification latérale :
Elle est caractérisée par le fait qu'elle s'effectue non seulement au sommet de la tige
mais également en arrière de celui-ci. La ramification latérale permet de distinguer
un axe principal et des axes latéraux. La plus grande partie de la ramification est
axillaire, c'est-à-dire que les rameaux naissent à l'aisselle des feuilles. Elle est
caractéristique des Spermaphytes, mais elle existe également chez les Ptéridophytes
(Prêles) et les Bryophytes.
Chez les Spermaphytes, les bourgeons sont insérés à l'aisselle d'une feuille ou d'une
bractée. On parle alors de bourgeons axillaires6
.
1 Cryptogame : En grec, caché et union ; globalement il s’agit des végétaux qui se reproduisent à l’aide de spores
2 Les Ptéridophytes sont un embranchement du règne des végétaux rassemblant les fougères
3Phanérogames : apparent et union Se dit des plantes dont les organes de reproduction sont apparents, se reproduisant par fleurs et par graines
4 Hyphaene est un genre de palmier qui comprend huit espèces. Hyphaene thebaica, du Soudan, fournit des fruits analogues aux dattes.
5 Chamaedorea est un genre de palmier de l'Amérique intertropicale comprenant de très nombreuses espèces.
6 Remarque : Un organe végétatif est qualifié d'adventif quand, s'ajoutant secondairement à d'autres organes du même type, il est d'une autre
origine et occupe une position différente. Ils sont à l'origine des rejets observés sur des troncs coupés (Carpinus betulus). Il n'existe pas de
rapport entre leur position et l'insertion des anciennes feuilles. Lorsqu'ils se forment sur la racine, ils sont à l'origine des drageons (Prunus
avium).

Sortie botanique du 16 février 2013
Site Natura 2000 de la reculée des Planches-Près-Arbois
Pascal Collin 2013 page 2
La ramification latérale est de deux types : monopodique ou sympodique.
• Les monopodes
Une tige monopodiale est caractérisée par un
bourgeon terminal fonctionnel d'une année
sur l'autre. La croissance de la tige s'effectue
par le bourgeon terminal. Ce type de
croissance est le modèle dominant, le Frêne
(Fraxinus excelsior) en est un bon exemple.
• Les sympodes
Une tige sympodiale est caractérisée par un
bourgeon terminal qui avorte ou qui porte une inflorescence. La tige est alors constituée par
une succession de pousses annuelles construites par des bourgeons différents. En fait, la
pousse annuelle est assurée par un bourgeon axillaire redressé. La croissance de type
sympodiale est plus rare que la croissance de type monopodiale. Elle existe particulièrement
chez le lilas et chez certains marronniers.
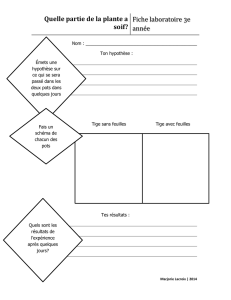
LA PHYLLOTAXIE
La phyllotaxie est une des nombreuses qualités héréditaires des plantes. Les feuilles
sont insérées sur les axes avec une grande régularité. Les causes de la phyllotaxie sont
liées aux propriétés du point végétatif (méristème). Diverses théories ont tenté de
rendre compte de ce phénomène. On retiendra :
la théorie classique qui admet l'existence d'une spire génératrice ou spire
principale. Elle est facilement visible sur les végétaux ligneux, il suffit de
compter le nombre de cicatrices foliaires superposées. Les droites par
Ci-dessus : coupe transversale dans un bourgeon de Chêne
rouge d’Amérique (Quercus rubra), F : feuilles, E :
écailles
Ci-contre :
En haut coupe longitudinale dans un embryon de Chêne
rouge d’Amérique (Quercus rubra), A : méristème, C :
cotylédons
En bas : coupe transversale dans un bourgeon de Frêne
commun (Fraxinus excelsior), F1 : paire de feuilles
opposées n°1, etc.

Sortie botanique du 16 février 2013
Site Natura 2000 de la reculée des Planches-Près-Arbois
Pascal Collin 2013 page 3
lesquelles passent ces superpositions sont appelées orthostiques. Des indices, ou
fractions phyllotaxiques, sont ainsi utilisés : 1/2 (beaucoup de monocotylédones), 1/3
(Cypéracées), 2/5 (beaucoup d'arbres), 3/8 (Verbascum), 5/13 (Picea), 8/21 (cônes
d'épicéa et de sapin), 13/24 (cônes de pin laricio). Chaque espèce a sa propre fraction.
Il est remarquable que les nombres qui constituent la fraction phyllotaxique
appartiennent à la suite infinie de Fibonacci7
La théorie des hélices foliaires est née des nombreuses anomalies ou lacunes
constatées dans la théorie classique (verticilles8
) ; les chercheurs ont élaboré une
théorie nouvelle dite des hélices foliaires. Cette théorie admet l'existence de plusieurs
hélices foliaires. Il existe cinq types de disposition des bourgeons, des feuilles et des
rameaux sur la tige : alterne distique, alterne hélicoïdale, opposée, opposée décussée et
verticillée.
7 L'apparition de la suite de Fibonacci est trop fréquente dans la nature pour être accidentelle. Elle permet de rendre compte de cette
croissance mécanique et automatique des feuilles (mais aussi des stipes des palmiers, des capitules de fleurs, des cônes, etc.). Cependant,
l'analyse d'une forme ne donne aucune explication sur sa génèse. La morphogénése de la plante et 'l'ontogénése' de la suite sont ignorées.
Pour une fraction phyllotaxique donnée il faut établir une explication de la croissance foliaire, capitulaire .... Il faut donc s'intéresser au
processus générique et par là même génétique de la croissance afin de répondre aux questions géométriques de la phyllotaxie. Alan Turing
(1912-1954) va chercher le principe mathématique qui permet de passer d'un état sans forme à celui de formes différenciées. Un modèle
différentiel nommé réaction-diffusion (RD) à partir duquel il parvient à montrer comment des processus physico-chimiques engendrent des
formes géométriques qui peuvent être de quelque aide pour l'interprétation des formes biologiques.
8 Lorsque les organes d’une plante sont insérés au même niveau, par groupe de trois unités au minimum, en cercle autour d'un axe (tige ou
rameau), on dit alors qu’ils sont en verticille ou verticillés.
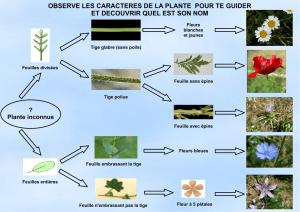
En haut :
. à gauche : Lamier Galéobdolon (Lamiastrum galeobdolon), feuilles opposées décussées.
. à droite : Camérisier noir (Lonicera nigra), feuilles opposées.
En bas :
. à gauche : Sceau de Salomon verticillé (Polygonatum verticillatum), feuilles en verticilles.
. à droite : Hêtre (Fagus sylvatica), feuilles alternes distiques.

Sortie botanique du 16 février 2013
Site Natura 2000 de la reculée des Planches-Près-Arbois
Pascal Collin 2013 page 4
Hêtre Président, forêt communale d’Arbois.
Hauteur de 35 m, circonférence de 3,15 m à 1m30 de haut, volume de 10 m
3
LA CROISSANCE
Dans la plupart des cas, l'allongement de la tige est plus
important que son accroissement diamétral. Seuls les bulbes
d'épicotyle de certaines crucifères (navet) présentent un
diamètre plus important que la hauteur de leur tige.
La croissance en longueur
Elle est très variable du point de vue de son intensité : de
quelque dm par an chez les pins à
60 cm/j chez certaines espèces de bambous. L'allongement
de la tige est différentiel car seul l'entrenœud s'allonge de
façon significative. On parle également de croissance
intercalaire. La croissance en longueur de la tige est en
principe indéfinie ; toutefois la taille des plantes est limitée.
La croissance en longueur est liée à l'existence des
méristèmes primaires situés à l'extrémité des tiges.
L'orientation de la tige sous l'action de l'attraction terrestre
présente un géotropisme négatif. Les branches qui sont
horizontales ou inclinées par rapport à la verticale présentent
une sensibilité moindre : il s'agit de diagéotropisme.
Au cours d'une année, l'allongement de la tige peut se faire
en une ou plusieurs fois. Dans ce dernier cas, on parle de
croissance rythmique ou de polycyclisme. Un grand nombre
de végétaux ligneux présentent une croissance rythmique
caractérisée par la succession de phases d'allongement rapide
et d'allongement lent ou nul de la tige. Durant la phase
d'allongement, la plante produit de nouvelles feuilles et de
nouveaux entre-nœuds pendant une durée qui varie selon les
espèces et les conditions de culture. Puis elle cesse de
s'allonger même si les conditions extérieures restent
favorables. Cet arrêt de l'allongement de la tige provoque
dans la plupart des cas l'apparition d'un bourgeon écailleux.
Après une durée variable selon les conditions de culture et
les espèces, le bourgeon débourre sans que les conditions de
culture aient varié.
Théophraste fut le premier à décrire les pousses successives
chez le Pin noir, le Sapin et le Chêne. Bien plus tard, de
nombreux travaux ont mis en évidence le caractère endogène
de la croissance rythmique d'un grand nombre de végétaux
ligneux de climat tempéré (Betula, Salix, Castanea,
Frangula, Corylus, Fagus, Quercus). C'est surtout le genre
Quercus qui a fait l'objet de nombreuses études. Ainsi
lorsqu'il est cultivé dès la germination en jours longs et à 25°
C, le Chêne pédonculé produit plusieurs vagues de croissance
dont la durée varie de 1 à 5 semaines. Quelques espèces
comme l'Alisier torminal présentent une croissance
rythmique atypique. Un temps de repos long (18 semaines)
sépare la première de la deuxième phase d'allongement de la

Sortie botanique du 16 février 2013
Site Natura 2000 de la reculée des Planches-Près-Arbois
Pascal Collin 2013 page 5
Le châtaignier aux cent chevaux, gouache de Jean-Pierre Houël
réalisée vers 1777.
tige. Enfin, d'autres espèces comme le Coudrier ou le Frêne commun manifestent durant leur
cycle de développement une rythmicité annuelle. Il existe donc une grande diversité dans
l'expression des rythmes de croissance chez les végétaux de climat tempéré.
La longueur de la tige est très variable et voici quelques records : Le Sapin pectiné (Abies
alba) Président de la forêt du Russey mesure 55 m pour 32 m3. Le sapin pectiné est le plus
grand arbre indigène d’Europe. Eucalyptus regnans : 132 m mesuré au théodolite en 1872 en
Australie, le plus grand arbre jamais mesuré. Doerner Fir est le nom d’un Douglas
(Pseudotsuga meinzies) de 100 m de haut, Oregon, USA. Hyperion est lui un séquoia à
feuilles d’if (Sequoia sempervirens) de Californie du nord est probablement l’arbre le plus
grand du monde en vie avec 115 m. Les arbres des forêts tropicales comportent de
nombreuses grandes espèces, les arbres de plus de 40 m sont fréquents9 et leur croissance
s’accélère encore aujourd’hui10
.
La croissance en diamètre
Elle se déroule en même temps que l'allongement de la tige et elle peut se prolonger après. La
croissance en diamètre trouve son origine dans l'existence de méristèmes secondaires ou
cambium. La croissance en diamètre n'est significative que chez les végétaux ligneux et
principalement les arbres.
Voici quelques records : le plus gros arbre du monde serait l’arbre de Tule au Mexique (image
page ci-après) avec plus de 50 m de circonférence. D’autres arbres tropicaux sont proches de
ce résultat, notamment les baobabs africains (Adansonia digitata) et les kauris géants néo-
zélandais (Agathis Australis). Par exemple, Te Matua Ngahere (père de la forêt en Maori) est
un Kauri de Nouvelle-Zélande dont la circonférence est de 16 m ; on estime son âge à 2 000
ans. Pour les baobabs, ce serait l’arbre de Platland ou de Sunland (Adamsonia digitata) en
Afrique du sud qui avec ses deux troncs connectés de plus de 10 m de diamètre chacun serait
le plus gros des baobabs (l’âge de cet arbre, sujet à controverse serait de 6 000 ans).
En Europe, il faut retenir le légendaire
châtaignier aux cent chevaux (Castanea
sativa) qui vivait sur les pentes de l’Etna et
qui avait une circonférence de 57,9 mètres
quand il a été mesuré en 1780. Abimé lors
d’intempéries, l’arbre est toujours vivant,
mais fragmenté.
En France, mis à part la tille d’Ivory (voir image ci-après), l’un des plus gros arbres de
l’hexagone serait un platane d'Orient (Platanus orientalis) situé à Fervaques dans le Calvados.
Son tronc présente une base conique, largement évasée, qui lui permet d’atteindre 12,5 m de
circonférence à 1m30 du sol. Mais là aussi, chauvinisme et esprit de clocher rendent parfois
les mesures plus ou moins élastiques…..
9 En Afrique, retenir le Moabi – Baillonella toxisperma, qui atteint 70 m de haut pour 5 m de diamètre…..en 600 ans environ ; en Amérique
tropicale, le noyer de l’Amazonie - Bertholletia excelsa - avec une hauteur maximum de l’ordre de 60 m ; et en Asie, le Méranti - Shorea
bracteolata – qui atteint là aussi 50 m de hauteur
10 Une équipe internationale de scientifiques a découvert que les arbres des forêts tropicales deviennent, en moyenne, plus grands. Ils
stockent plus de carbone de l’atmosphère dans leurs troncs. Les arbres des forêts tropicales intactes absorbent presque 20% des émissions de
CO2 dues aux combustibles fossiles. Les chercheurs démontrent que les forêts tropicales absorbent un chiffre étonnant de 4,8 milliards de
tonnes (ou GT) de CO2 de l’atmosphère par an. Les émissions mondiales représenteraient environ 35 GT par an. Article publié dans le
journal Nature : n° 457, 1003-1006, 19 February 2009.
 6
7
8
9
6
7
8
9
1
/
9
100%



![Morphologie des Angiospermes [Mode de compatibilité]](http://s1.studylibfr.com/store/data/000565368_1-a68c29846ff57d73904763e33ee955b5-300x300.png)