Implication du régulateur global CsrA dans le maintien de l

Implication du régulateur global CsrA dans le maintien de
l’homéostasie de l’enveloppe chez Escherichia coli
Mémoire de Thibaut HALLAERT, 2012-2013
Présentation dans le cadre du concours de l’A.Sc.Br
J’ai réalisé mon mémoire au sein du laboratoire de génétique et physiologie bactérienne (LGPB) qui
fait partie de l’Institut de Biologie et de Médecine Moléculaire (IBMM) de l’Université Libre de
Bruxelles (ULB), sous la supervision du professeur Laurence Van Melderen et d’Ethel Seyll, en vue de
l’obtention du diplôme de Master en biochimie et biologie moléculaire et cellulaire.
Les bactéries vivent dans des environnements en perpétuel changement. Pour s’y développer de
manière efficace, elles doivent être capables de percevoir l’état du milieu et de répondre à toutes les
modifications pouvant y survenir, tout en tenant compte de leur propre état. L’ensemble de ces
informations sont générées et traitées par un réseau complexe de voies de signalisation dont l’étude
appartient au domaine de la physiologie bactérienne.
Mises en place et constamment raffinées par l’évolution, ces voies de signalisation sont composées
de récepteurs sensibles aux signaux activant des cascades de réactions convergeant vers les
régulateurs globaux. Ces derniers régulent des fonctions très diverses de manière cohérente pour
rendre la bactérie plus apte à se développer dans l’environnement ayant produit le signal initial. Par
conséquent, les régulateurs globaux jouent un rôle de contrôle central en physiologie bactérienne.
Au LGPB, nous étudions le régulateur global CsrA (Carbon Storage Regulator A) modulant des
fonctions métaboliques et des comportements sociaux chez de nombreuses espèces bactériennes.
Sous sa forme active, CsrA régule positivement la consommation des sources de carbone pour
fournir de l’énergie à la bactérie (voie de la glycolyse et du cycle de Krebs) et négativement leurs
mises en réserve (voie de la glycogénogenèse). De plus, il inhibe la production d’exopolysaccharide
permettant aux bactéries d’adhérer aux surfaces et favorise la mise en place du flagelle, un outil
locomoteur impliqué dans le mouvement natatoire. Enfin, il réprime l’expression du senseur du
quorum et régule positivement l’expression de facteurs de virulence. CsrA est ainsi impliqué dans le
switch entre les deux modes de vie bactériens principaux, le mode ‘solitaire, mobile et virulent’, qu’il
active et le mode ‘communautaire, sessile et persistant’, qu’il réprime. Ce second mode de vie est
caractérisé par la formation de colonies multicellulaires adhérant aux surfaces et protégées des
agressions environnementales (prédateurs, antibiotiques, système immunitaire,…) par une matrice
extracellulaire complexe. Ces ‘biofilms’ sont responsables de problèmes sanitaires et médicaux tels
que la chronicité de certaines infections bactériennes. CsrA est inactivé lorsque la concentration en

déchets métaboliques (acétate, formate) augmente dans le milieu indiquant à la fois une grande
densité de population et un appauvrissement de l’environnement.
Nous avons supprimé le gène csrA du génome d’une souche bactérienne d’Escherichia coli
uropathogène. Ces bactéries mutantes, dépourvues du régulateur, sont hyper-adhérentes et forment
des réserves énergétiques conformément à nos connaissances sur les fonctions régulées par CsrA. Au
cours de mon mémoire, nous avons découvert qu’en plus de ces caractéristiques attendues, les
bactéries mutantes souffrent d’altérations de leur enveloppe.
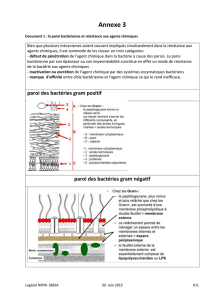
Comme toute cellule vivante, les cellules bactériennes sont délimitées par une membrane composée
de lipides et de protéines s’organisant en une mosaïque fluide. A l’extérieur de celle-ci, les bactéries
disposent d’un exosquelette rigide et, dans le cas de la famille de bactéries que nous étudions (les
gram-négatives), d’une seconde membrane composée d’un type de lipides particulier formant un
bouclier dense et figé, traversé de pores protéiques à sélectivité variable. L’ensemble de ces
structures extra-cytoplasmiques représente l’enveloppe et constitue l’interface entre
l’environnement et la bactérie. L’enveloppe assure les fonctions de barrière, de frontière d’échanges
(acquisition de nutriments et d’informations, excrétion de déchets, sécrétion de composés) mais est
aussi impliquée dans des processus plus fondamentaux tels que la production d’énergie.
En découvrant que les bactéries dépourvues de CsrA voient leur enveloppe affectée, nous ouvrons
deux grandes perspectives dans le domaine de la physiologie bactérienne. Soit ces altérations de
l’enveloppe sont une conséquence directe de l’absence de CsrA indiquant une nouvelle fonction du
régulateur en lien avec l’homéostasie de ces structures, soit il s’agit de conséquences indirectes dues
aux perturbations métaboliques que l’absence de CsrA engendre, mettant en évidence des
interactions entre le métabolisme énergétique et les voies de synthèses de l’enveloppe.
Ces hypothèses seront mises à l’épreuve dans le cadre d’une thèse de doctorat. De nombreuses et
passionnantes questions restent ouvertes à propos du fonctionnement et de l’origine évolutive de
l’enveloppe bactérienne. De plus, de part l’implication de CsrA dans les processus infectieux et dans
la gestion des flux de carbone chez les bactéries, les découvertes associées à son étude pourraient
avoir un impact dans les domaines médicaux et de la bioingénierie (production de biofuel, cultures
bactériennes, etc.).
1
/
2
100%