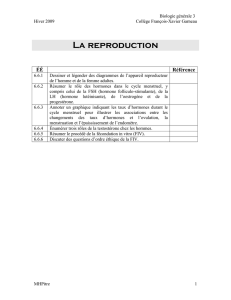
TABLE DES MATIÈRES - Papyrus : Université de Montréal

Université de Montréal
Étude sur l’oxygénation des lits capillaires du disque
optique au cours du cycle menstruel chez les femmes
par
Jessy Hilal
École d’optométrie
Mémoire présenté à la Faculté des études supérieures
en vue de l’obtention du grade de maîtrise
en sciences de la vision
Décembre, 2011
© Jessy Hilal, 2011

ii
Université de Montréal
Faculté des études supérieures
Ce mémoire intitulé :
Étude sur l’oxygénation des lits capillaires du disque optique au cours du cycle
menstruel chez les femmes
présenté par :
Jessy Hilal
a été évalué par un jury composé des personnes suivantes :
Jean-François Bouchard, président-rapporteur
Vasile Diaconu, directeur de recherche
Elvire Vaucher, membre du jury

iii
Résumé
Il est connu qu’on retrouve chez les femmes en post-ménopause un risque plus important
de développer des maladies oculaires comparativement aux hommes du même groupe
d’âge. Il semble que les changements hormonaux, et en particulier la baisse importante des
niveaux d’estradiol, secondaires à la sénescence folliculaire constituent un facteur
étiologique à long terme. Cela étant, il est légitime de se demander si les variations des
niveaux d’hormones sexuelles endogènes peuvent également occasionner des effets à court
terme sur les tissus de l’œil. Cette interrogation constitue d’ailleurs le motif principal de
l’élaboration de la présente étude.
Sachant qu’il se produit chez les femmes non ménopausées des variations continuelles des
niveaux d’hormones sexuelles stéroïdiennes au cours de leur cycle menstruel, des femmes
en âge de procréer ont été recrutées comme sujets d’étude. Dans un deuxième temps, afin
de trouver le paramètre d’intérêt, on a effectué une revue de la documentation scientifique
qui révèle un fait bien établi : les estrogènes favorisent la vasodilatation des vaisseaux
sanguins par l’intermédiaire du monoxyde d’azote, et permettent, par le fait même,
l’accroissement du débit sanguin tissulaire. Or, comment mesurer des variations de débit
sanguin dans des tissus oculaires? Comme il est expliqué dans la discussion du présent
mémoire, les variations d’oxygénation dans un organe dont le métabolisme est
relativement stable sont le reflet de variations de débit sanguin. Grâce à une technique de
mesure basée sur la spectroréflectométrie, il est possible de mesurer le taux
d’oxyhémoglobine (HbO2) des lits capillaires du disque optique. En observant les

iv
variations du taux d’oxyhémoglobine au cours du cycle menstruel chez les sujets, on peut
ainsi mesurer l’effet des variations hormonales cycliques sur l’irrigation des tissus
oculaires.
En somme, l’objectif de cette recherche est de mieux comprendre, en suivant le cycle
menstruel des femmes, l’effet des hormones sexuelles endogènes sur l’oxygénation des lits
capillaires du disque optique. Étant à la base du métabolisme de l’œil, l’apport en oxygène
et en divers substrat véhiculés par la circulation sanguine est important au maintien de la
santé oculaire. L’éclaircissement du lien entre les hormones et l’oxygénation de la rétine
constituerait un avancement important, puisqu’il permettrait de comprendre pourquoi
certaines atteintes oculaires, comme la cécité, touchent davantage les femmes.
Les résultats de cette étude ont démontré que le taux d’oxyhémoglobine mesuré dans les
lits capillaires du disque optique de l’œil ne subit pratiquement pas de variations
significatives durant le cycle menstruel lorsqu’on considère les incertitudes des valeurs
mesurées. Également, on observe une variabilité similaire des taux d’oxyhémoglobine
mesurés chez les femmes en âge de procréation et chez les hommes du même groupe
d’âge. Cela suggère que les changements hormonaux cycliques, qui ne se produisent que
chez les femmes, n’occasionnent probablement pas de variation significative mesurable du
taux d’oxyhémoglobine. Bref, malgré les effets possibles des estrogènes sur le diamètre
artériolaire, il semble que les mécanismes locaux de régulation du débit sanguin tissulaire
maintiennent un état d’équilibre propre au tissu irrigué et adapté aux besoins métaboliques
locaux.

v
Mots clés : oxyhémoglobine, oxygénation du nerf optique, maladies oculaires, hormones
sexuelles stéroïdiennes, cycle menstruel.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
1
/
152
100%