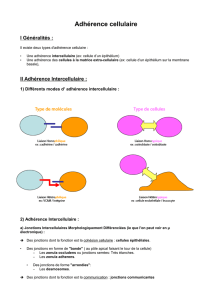
Les dispositifs d`adhérence jouent des rôles fondamentaux dans le

1
Devoir N°9 2
ème
partie
17/06/09
BIOLOGIE
Épreuve B
Durée : 2 heures 30 minutes
______
L’usage de la calculatrice, d’abaques et de tables est interdit pour cette épreuve.
Si, au cours de l’épreuve, un candidat repère ce qui lui semble être une erreur d’énoncé, il le signale
sur sa copie et poursuit sa composition en expliquant les raisons des initiatives qu’il a été amené à prendre.
Les dispositifs d’adhérence jouent des rôles fondamentaux dans le
déroulement du développement embryonnaire.
A partir de l’exploitation des documents et de vos connaissances, mettez en
évidence quelques uns de ces rôles
* L’exposé sera encadré par une introduction et une conclusion et sera structuré par un
plan faisant apparaître explicitement les thèmes abordés et la progression suivie.
* L’exposé doit se limiter aux trois thèmes abordés par les documents, qui font l’objet de
trois parties sont indépendantes.
* Les documents peuvent être découpés et collés sur la copie à condition d’être légendés,
commentés ou exploités.
Les documents proposés portent
- pour la partie I, sur le creusement de la cavité amniotique chez les mammifères
- pour la partie II, sur la formation du tube neural chez les amphibiens
- pour la partie III, sur la morphogenèse chez la drosophile

2
THEME 1 : La formation de la cavité amniotique au cours du développement embryonnaire des
Mammifères.
Document 1-1 : L’étape d’amniogenèse
Pendant le développement embryonnaire des mammifères, se forme une cavité qui deviendra la cavité
amniotique (= « poche des eaux »).
Coupe de blastocyste à l’issue de l’amniogenèse
Chez les primates, la formation de cette cavité ou amniogenèse s’effectue par cavitation. La cavitation est la
mort des cellules centrales ectodermiques sous l’action d’une molécule-signal provenant des cellules
endodermiques.
Seule une couche de cellules ectodermiques survit. A la base de cette couche, on trouve une lame basale très
riche en fibronectine.
In vitro, isolées de l'embryon, toutes les cellules ectodermiques, mises en contact avec la molécule-signal
provenant de l’endoderme meurent, y compris les cellules isolées à partir de la couche externe de
l'ectoderme.
Document 1-2 : Rôle de la fibronectine sur la survie de fibroblastes
Afin de comprendre les causes de survie des cellules ectodermiques entourant la cavité amniotique, des
expérimentations sont menées. Les cellules prises pour modèles expérimentaux sont ici des fibroblastes.
Des molécules de
fibronectine
(glycoprotéines) sont
fixées sur une lame de
verre de la façon
présentée ci-contre.
Dans les deux cas, la
même quantité totale de
fibronectine est fixée sur
la lame de verre.
Des cellules de tissu
conjonctif (fibroblastes)
sont mises en culture sur
cette lame. On observe les
cellules cultivées au
microscope.
Quantité X de
fibronectine fixée
en un unique spot
Quantité X de
fibronectine fixée
en plusieurs spots
différents
Les
cellules
meurent par
apoptose
Les
cellules
vivent,
grandissent
et se
multiplient

3
Document 1-3 : Intervention des intégrines dans la survie des cellules
L’étude se poursuit sur d’autres cellules, des hépatocytes (cellules du foie).
Des hépatocytes sont mis en culture in vitro sur des
matrices extracellulaires artificielles (constituées
principalement de collagène I et de fibronectine).
Après une semaine de culture, les cellules ont
proliféré et présentent un phénotype normal.
On ajoute alors à la culture :
- soit rien (contrôle : ctrl)
- soit des anticorps spécifiques des intégrines
membranaires, qui se lient et recouvrent les sites de
fixation de ces intégrines aux constituants matriciels
(αvβ3) ;
On mesure alors dans les cultures le taux de mort
cellulaire programmée des cellules (index
d'apoptose).

4
THEME 2 : La formation du tube neural chez les Amphibiens
Documents extraits de Biologie du développement Wolpert (Ed.Dunod)
Document 2-1 : Culture de cellules de neurectoderme et d’épiderme de neurula
Des explants de neurulas (stade plaque neurale) sont
prélevés au niveau de l'épiderme d'une part, et au niveau
du neurectoderme d'autre part.
Les cellules des explants sont dissociées et mises en
culture ensemble dans une même boîte de Pétri.
Les cellules s'agrègent les unes aux autres, dans une
configuration particulière représentée dans la 3
e
partie du
document.
Document 2-2 : Cadhérines et formation du tube neural
A: La cadhérine E, la cadhérine N et la protéine N-CAM sont localisées au niveau de la région dorsale de l'embryon
au cours de la neurulation.
B: Une expression anormale du gène codant la cadhérine N est déclenchée dans l'embryon. La figure présente le profil
d'expression du gène CAD N de la cadhérine N et le résultat de la neurulation dans cet embryon.

5
THEME 3 : Adhérence différentielle et morphogenèse
D’après Médecine Science Mars 2007 article d’ I. Bécam
Document 3-1 : Ségrégation et tension superficielle des cellules
Dans les années 1960, Malcom Steinberg propose l’hypothèse d’adhérence différentielle : selon cette
hypothèse, les tissus embryonnaires sont assimilés à des liquides et les lois physiques qui gouvernent le
comportement de deux liquides, l’eau et l’huile par exemple, dirigeraient aussi les réarrangements
tissulaires. Un des principes fondamentaux de la thermodynamique prédit qu’un système tend toujours vers
l’état le plus stable, ce qui se mesure par une décroissance de son énergie libre. Dans le cas de deux liquides,
une partie de l’énergie libre provient des forces de tension qui s’exercent aux surfaces de contact. Cette
énergie libre est d’autant plus grande que les forces de tension de surface sont importantes et la surface de
contact étendue. Si la tension de surface est importante, l’énergie libre peut être minimisée en réduisant la
surface de contact. Dans l’exemple de l’émulsion eau-huile, l’huile, dont la tension de surface est plus
importante que celle de l’eau, forme de petites gouttes sphériques (plus petit rapport surface/volume)
entourées d’eau.
Pour démontrer que ces lois physiques s’appliquent aux tissus, l’équipe de M. Steinberg mesure la tension
de surface au sein des différents agrégats cellulaires de cellules dissociées à partir de différents tissus puis
observe l’organisation spontanée d’un agrégat comprenant des cellules provenant de 2 tissus différents.
Plus récemment, en 2005, M. Steinberg montre dans un système de culture cellulaire in vitro que la tension
de surface tissulaire est directement proportionnelle à la quantité de cadhérines exprimées par les cellules.
Rouge
vert
jaune
rouge
bleu
jaune
orange
bleu
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%