Reconnaissance innée Reconnaissance des

Reconnaissance innée
Dr. Isabelle CREMER
MCU Université Paris 6
U872 INSERM, équipe 13: « Microenvironnement Immunitaire et Tumeurs »
Centre de Recherche des Cordeliers. 75006 Paris
Reconnaissance des microorganismes par
les molécules et les cellules du système
immunitaire

Plan du cours
Barrières anatomiques
Barrières physiologiques
Reconnaissance par les cellules du système immunitaire:
Les cellules tueuses naturelles NK, lymphocytes Tγδ
Les cellules phagocytaires
Reconnaissance par les molécules du système immunitaire:
Les récepteurs Toll-like « TLR » !
Les lectines de type C
Les récepteurs « scavengers » (poubelles)
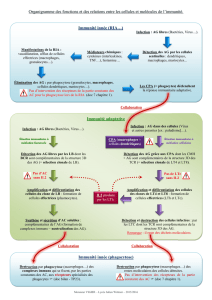
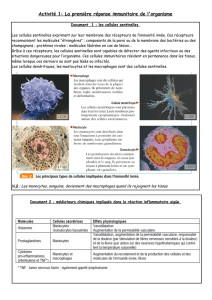
Introduction

Différentes voies d’entrée des pathogènes
Immunobiology (Janeway
et al
.)

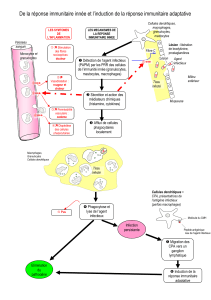
Réponse à une première infection: 3 phases
Immunobiology (Janeway
et al
.)
Immunité adaptative plus tardive. Temps d’expansion clonale des LT et LB spécifiques du
pathogène.

Comparaison entre immunité innée et adaptative
D’après McGuinness D.H.
et al
. 2003. Trends in Parsitology 19:312
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
1
/
43
100%