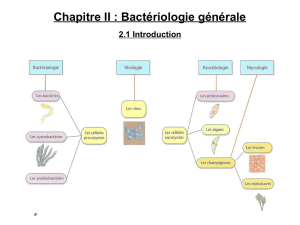
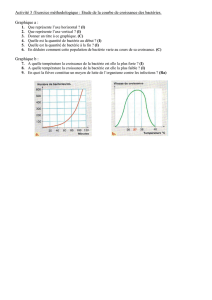
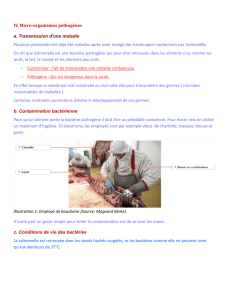
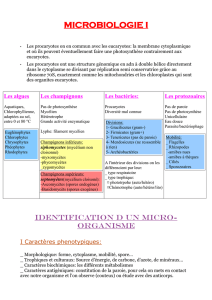
Evaluation des propriétés immunomodulatrices de la bactérie

UNIVERSITÉ LIBRE DE BRUXELLES
Ecole Interfacultaire de Bioingénieurs
Laboratoire d’Allergologie Expérimentale
Evaluation des propriétés immunomodulatrices de la
bactérie lactique Lactobacillus plantarum NCIMB8826
dans le cadre de l'allergie aux acariens
Thèse présentée en vue de l’obtention du grade de
Docteur en Sciences Agronomiques et Ingénierie
Biologique
Peter Rigaux
2008
Promoteur : Alain Jacquet
Co-promoteur : Muriel Moser


UNIVERSITÉ LIBRE DE BRUXELLES
Ecole Interfacultaire de Bioingénieurs
Laboratoire d’Allergologie Expérimentale
Evaluation des propriétés immunomodulatrices de la
bactérie lactique Lactobacillus plantarum NCIMB8826
dans le cadre de l'allergie aux acariens
Thèse présentée en vue de l’obtention du grade de
Docteur en Sciences Agronomiques et Ingénierie
Biologique
Peter Rigaux
2008
Promoteur : Alain Jacquet
Co-promoteur : Muriel Moser

Je tiens à remercier la Commission Européenne, le Fond pour la formation à la Recherche
dans l’Industrie et dans l’Agriculture (FRIA) et la Fondation Alice et David Van Buuren qui m’ont
permis à travers leur programme de financement, de réaliser mes cinq années de thèse.
Je remercie chaleureusement le Dr. Alain Jacquet, mon promoteur de thèse, de m’avoir
accueilli au sein de son laboratoire, de m’avoir soutenu et guidé pendant toutes ces années.Merci à
toute l’équipe du laboratoire d’Allergologie Expérimentale : Lida (une aide très précieuse), Louis pour
tes différents coups de main et pour ton expertise technique, Manu (qu’ils sont beaux ces Western
Blot !), Jonathan pour ton aide et tes conseils, Virginie pour ton aide et ta disponibilité, les étudiants
de passage (Farshid, Céline C., Nicolas, Laurence) pour leur aide même si elle a parfois été ponctuée
de quelques sueurs froides et tous les autres membres du laboratoire qui ont su y apporter une
ambiance chaleureuse (Jean-François, Céline M., Céline B., David, Christelle, Virginie, Eugénie,
Mauro, les inoubliables « Quoiquois » et Delphine)
Je remercie le Dr. Bruno Pot de m’avoir accueilli dans son laboratoire. Je tiens à exprimer ma
plus vive gratitude au Dr. Catherine Daniel pour son aide, sa gentillesse, ses conseils et sa
disponibilité. Je remercie Michael Hisbergues pour le travail essentiel qu’il a fourni. Merci à tout le
reste de l’équipe et en particulier à Sabine. Je remercie aussi nos colocataires pour leur convivialité.
Je remercie vivement le Dr. Joel Pestel de m’avoir accueilli au sein de l’unité U416, d’avoir
initié ce projet de collaboration, de m’avoir conseillé et de m’avoir soutenu. Je remercie également
toute l’équipe de l’unité U416 et particulièrement Céline Ratajczak et Philippe Marquillies.
Je remercie le Dr. Muriel Moser d’avoir accepté d’être la co-promotrice de cette thèse.
Enfin, last but not least, je remercie ma famille pour m'avoir apporté un soutien affectif et
matériel pendant toute cette aventure.

ADN : Acide DésoxyriboNucléique
APC : Antigen Presenting Cell
BMDC : Bone Marrow-derived Dendritic Cell
CD : Cluster of Differentiation
CMH : Complexe Majeur d’Histocompatibilité
CFSE : CarboxyFluoresceine Succinimidyl Ester
CpG : deoxy-Cytidylate-phosphate-deoxyGuanylate
DC : Dendritic Cell
DC-SIGN : Dendritic Cell-Specific Intercellular adhesion molecule-3 Grabbing Nonintegrin
ERK : Extracellular signal-Regulated Kinase
GM-CSF : Granulocyte Macrophage Colony Stimulating Factor
IFN : InterFéroN
IL : InterLeukin
JNK : c-Jun N-terminal Kinase
KO : Knock Out
LPS : LipoPolySaccharide
LTA : LipoTeichoic Acid
MAPK : Mitogen-Activated Protein Kinase
MCP : Monocyte Chemotactic Protein
MD-DC : Monocyte Derived Dendritic Cell
MIP : Macrophage Inflammatory Protein
MR : Mannose Receptor
MyD88 : Myeloblast Differentiation factor 88
NK : Natural Killer
ODN : OligoDeoxyNucleotide
OVA : OVAlbumin
PAMPs : Pathogen Associated Molecular Patterns
PBMC : Peripheral blood mononuclear cells
PG : ProstaGlandin
PGN : PeptoGlycaN
PRRs : Pattern Recognition Receptors
RANTES : Regulated upon Activation, Normal T-cell Expressed, and Secreted
STAT : Signal Transducer and Activator of Transcription
TcR : T cell Receptor
TGF : Tranforming Growth Factor
TH: T Helper cell
TIR : Toll/Interleukin-1 Receptor
TLR : Toll Like Receptor
TNF : Tumor Necrosis Factor
WT : Wild Type
ZO-1 : Zonula Occludens 1
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
1
/
108
100%