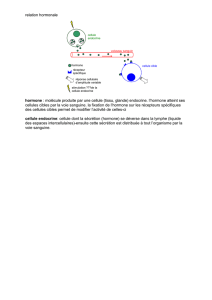
La structure des récepteurs de la prolactine et de l`hormone de

1990 6 : 778-84
Paul A. Kelly
Jean Dj iane
Jean-Marie Boutin
Marc Edery
La structure des récepteurs de
la prolactine
et de l'hormone de croissance
est maintenant connue
L


A

1
l

°
 6
7
6
7
1
/
7
100%