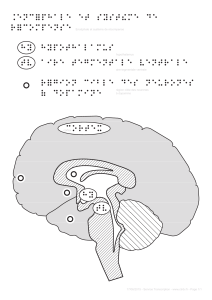
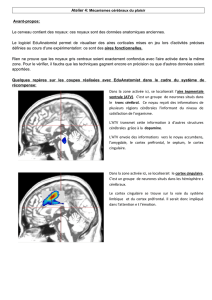
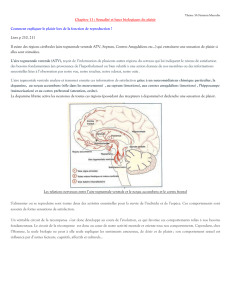
Le cortex préfrontal et la dopamine striatale dans l`apprentissage

Le cortex pr´efrontal et la dopamine striatale dans
l’apprentissage guid´e par la r´ecompense : conception et
´etude d’une tˆache cognitive d’exploration par essais et
erreurs en imagerie par r´esonance magn´etique
fonctionnelle et en tomographie par ´emission de positons
avec le 11C-raclopride
Claire Landmann
To cite this version:
Claire Landmann. Le cortex pr´efrontal et la dopamine striatale dans l’apprentissage guid´e par
la r´ecompense : conception et ´etude d’une tˆache cognitive d’exploration par essais et erreurs en
imagerie par r´esonance magn´etique fonctionnelle et en tomographie par ´emission de positons
avec le 11C-raclopride. Neurosciences [q-bio.NC]. Universit´e Pierre et Marie Curie - Paris VI,
2007. Fran¸cais. <tel-00160868>
HAL Id: tel-00160868
https://tel.archives-ouvertes.fr/tel-00160868
Submitted on 9 Jul 2007
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-
entific research documents, whether they are pub-
lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destin´ee au d´epˆot et `a la diffusion de documents
scientifiques de niveau recherche, publi´es ou non,
´emanant des ´etablissements d’enseignement et de
recherche fran¸cais ou ´etrangers, des laboratoires
publics ou priv´es.


THESE de DOCTORAT de L’UNIVERSITE PARIS 6
Spécialité : NEUROSCIENCES COGNITIVES
Présentée par Claire LANDMANN
pour l’obtention du titre de Docteur de l’Université Paris 6
LE CORTEX PREFRONTAL ET LA DOPAMINE
STRIATALE DANS L’APPRENTISSAGE
GUIDE PAR LA RECOMPENSE.
Conception et étude d’une tâche cognitive d’exploration par essais et
erreurs en imagerie par résonance magnétique fonctionnelle et en
tomographie par émission de positons avec le 11C-raclopride.
Thèse effectuée sous la direction du Pr. Stanislas DEHAENE
et la codirection du Dr. Sabina PAPPATA
dans l’unité INSERM 562, SHFJ/CEA/DRM/DSV
Soutenue le 26 juin 2007 devant le jury composé de :
Dr. Jean-Claude Dreher Rapporteur
Pr. Eric Salmon Rapporteur
Dr. Etienne Koechlin Examinateur
Pr. Laurent Cohen Examinateur

2

3
Remerciements
Le volet de psychophysique et d’IRMf de ce travail s’est déroulé sous la direction de Stanislas
Dehaene, que je tiens à remercier pour son encadrement scientifique, son enthousiasme communicatif
et son soutien. J’ai énormément appris, et pris beaucoup de plaisir à travailler au sein de son équipe. Je
tiens également à remercier Sabina Pappata, qui a encadré le volet de TEP de ma thèse, pour son
encadrement scientifique, sa rigueur et sa disponibilité.
Je remercie également :
Jean-Claude Dreher, Eric Salmon, Etienne Koechlin et Laurent Cohen qui me font l’honneur et le
plaisir de participer au jury de ma thèse,
Sébastien Mériaux et Marie-Claude Grégoire pour leur importante contribution au développement des
outils d’analyse et au traitement des données de TEP,
Antoinette Jobert pour son aide dans les différentes étapes de traitement des images,
Mariano Sigman pour son aide à tous les niveaux, son soutien précieux, son ouverture d’esprit, …
Christophe Pallier, pour sa grande disponibilité et son aide au « domptage » de toutes espèces de
programmes informatiques,
Andreas Kleinschmidt pour ses conseils et sa lecture attentive de l’article d’IRMf,
Avec eux, les autres membres de l’équipe pour l’environnement gai et stimulant qu’ils ont créé, en
particulier Teodora Gliga, Véronique Izard, Claire Sergent, et Philippe Pinel, ainsi que Ghislaine
Dehaene-Lambertz, Ed Hubbard, Manuella Piazza, Fabien Vinckier, Antoine Del Cul, Marco Buiatti,
Laurent Cohen, Lionel Naccache, Jérôme Sackur, Joachim Forget, Anne-Dominique Devauchelle,
Marie-Hélène Baju,
Denis Rivière, Alexis Roche, Yann Cointepas, Philippe Ciuciu, Arnaud Cachia, Edouard Duchesnay,
Renaud Maroy, et tous les membres de l’Unité de neuroimagerie anatomo-fonctionnelle (UNAF) pour
le développement d’outils de visualisation et d’analyse des images, et pour leur aide et leur
disponibilité,
Michel Bottlaender et Dimitri Roumenov qui ont réalisé l’examen médical des sujets de TEP ; les
infirmiers, et les membres du Groupe d’acquisition et de traitement de l’image (GATI), en particulier
Laurence Laurier, Sandrine Bourgeois, Patrick Bodilis, et Vincent Brulon, qui ont rendu très agréables
les heures d’acquisition des images de TEP,
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
1
/
215
100%