TABLE DES MATIÈRES

Table des Matières - 1
Département de biologie, Université d’Ottawa
TABLE DES MATIÈRES
TABLE DES MATIÈRES 1
INTRODUCTION 3
La diversité animale 3
Besoins des animaux 4
Oxygène 4
Eau 5
Nourriture 5
Reproduction 5
Fonctions nécessaires à la vie animale 5
Circulation 6
Excrétion 6
Locomotion 6
Perception 6
Coordination 6
Contraintes liées à l'environnement 7
Le milieu marin 7
Le milieu d'eau douce 7
Le milieu terrestre 7
CLASSIFICATION ET ÉVOLUTION ANIMALE 9
Taxonomie (ou taxinomie) 9
Classification hiérarchique 10
Approche traditionnelle 10
Phénétique 11
Cladistique 11
Les cinq Règnes 12
Grandes divisions phylogénétiques du règne animal
12
Uni- versus Pluricellulaires 12
Agrégats de cellules versus tissus 13
Plan de symétrie 13
Cavité interne 13
Protostomiens vs Deutérostomiens 13
Échelle temporelle 13
Précambrien (4600-570M) 14
Paléozoïque (570-230M) 14
Mésozoïque (230-65M) 14
Cénozoïque (64M-Aujourd'hui) 14
LES PROTOZOAIRES 15
Locomotion et support 17
Respiration et circulation 18
Alimentation et digestion 19
Excrétion et osmorégulation 19
Reproduction 20
Défenses 22
Écologie 22
LES PORIFÈRES 25
Généralités 25
Architecture et classification 26
Respiration et circulation 28
Alimentation et digestion 28
Excrétion et osmorégulation 28
Reproduction 28
Défenses 29
Écologie 30
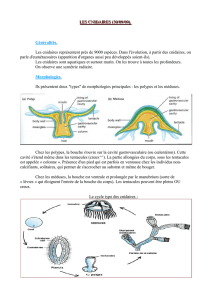
LES CNIDAIRES 31
Architecture et classification 32
Locomotion et support 33
Respiration et circulation 34
Alimentation et digestion 35
Excrétion et osmorégulation 36
Sens et système nerveux 36
Reproduction 36
Défenses 37
Écologie 37
LES PLATHELMINTHES 39
Origine des métazoaires et des animaux bilatéraux
40
Architecture et classification 40
Locomotion 42
Respiration et circulation 42
Alimentation et digestion 42
Excrétion et osmorégulation 42
Reproduction 43
Sens et système nerveux 44
Défenses et adaptations 44
LES NÉMATODES 47
Architecture 47
Locomotion 48
Respiration et circulation 48
Alimentation et digestion 48
Excrétion 49
Reproduction 49
Écologie 49
LES MOLLUSQUES 51
Architecture et classification 51
Locomotion 54
Respiration et circulation 54
Alimentation et digestion 55
Reproduction 56
Défenses 56

2 - Animaux: Structures et Fonctions
BIO 2525 - Hiver 2003
Écologie 56
LES ANNÉLIDES 57
Architecture et Classification 58
Locomotion 59
Respiration et circulation 61
Alimentation et digestion 61
Excrétion et osmorégulation 62
Système nerveux 62
Reproduction 62
Une parenthèse sur la reproduction 63
Reproduction sexuée ou asexuée? 63
Si sexuée, monoïque ou dioïque? 63
Qui fait quoi? 64
Écologie 64
LES ARTHROPODES 65
Architecture et classification 65
Locomotion 66
Respiration et circulation 67
Alimentation et digestion 69
Excrétion et osmorégulation 69
Cycle biologique 70
Défenses 70
Écologie 70
LES ÉCHINODERMES 73
Architecture et classification 74
Locomotion 75
Respiration et circulation 75
Alimentation et digestion 75
Excrétion et osmorégulation 76
Sens et système nerveux 76
Reproduction 76
Défenses et adaptations à la vie sessile 76
Écologie 77
INTRODUCTION AUX CHORDÉS 79
Architecture et classification 80
Origine des Chordés 81
Évolution des Chordés 82
Les Vertébrés 82
LES POISSONS 83
Architecture et classification 84
Locomotion 85
Respiration et circulation 85
Osmorégulation et excrétion 87
Sens et système nerveux 87
Écologie 87
LES AMPHIBIENS 89
Adaptations des Vertébrés à la vie terrestre 89
Architecture et classification 91
Squelette et locomotion 91
Respiration et circulation 91
Alimentation et digestion 92
Excrétion et osmorégulation 92
Reproduction 93
LES REPTILES 95
Architecture et Évolution 95
Respiration et circulation 96
Alimentation et digestion 97
Excrétion et osmorégulation 97
Reproduction 97
Défenses et adaptations 98
Système nerveux 98
LES OISEAUX 99
Modifications associées au vol 99
Respiration et circulation 100
Alimentation et digestion 101
Excrétion et osmorégulation 101
Système nerveux 101
LES MAMMIFÈRES 103
Architecture et classification 103
Locomotion 104
Respiration et circulation 104
Alimentation et digestion 105
Excrétion et osmorégulation 105
Système nerveux 105
Adaptations 106
GLOSSAIRE 107

Introduction - 3
Département de biologie, Université d’Ottawa
Introduction
La diversité des animaux est incroyable. En taille, les animaux peuvent
varier de 0.1 mm à quelques dizaines de mètres. Certains ne vivent que quel-
ques jours, d’autres plus d’un siècle. On retrouve des animaux dans tous les
habitats, allant des fosses abyssales aux pics des plus hautes montagnes.
Pourtant, malgré leurs nombreuses différences, tous ces organismes ont plu-
sieurs choses en commun. Ils proviennent vraisemblablement d’un même
ancêtre, ont fondamentalement les mêmes besoins, et font face aux mêmes
contraintes imposées par le milieu dans lequel ils vivent.
La diversité animale
Il y a présentement environ 1,500,000 espèces animales qui sont con-
nues. On soupçonne que les espèces vivant actuellement, mais qui n'ont pas
encore été décrites, sont au moins aussi nombreuses, sinon de cinq à trente
fois plus nombreuses que les espèces connues. Le nombre d’espèces qui ont
vécu sur la terre mais qui ont maintenant disparu est fort probablement
beaucoup plus grand que celui des espèces vivant aujourd’hui.
Les zoologistes regroupent les animaux multicellulaires en 32 embran-
chements. Chaque embranchement est caractérisé par une architecture parti-
culière et une série de propriétés biologiques qui le distingue de tous les
autres. Presque tous ces embranchements sont les survivants d’un groupe
plus grand de près d’une centaine d’embranchements qui existaient sur Terre
il y a 600 millions d’années lors du Cambrien. Il s’est alors produit une
grande diversification de la faune, sans doute l’évènement évolutionnaire
le plus important de l’histoire de la vie animale. Au cours d’une période de
quelques millions d’années, toutes les architectures animales existant
aujourd’hui, et une multitude maintenant disparues et connues seulement
par les fossiles, sont apparues.
Le nombre d'espèces dans chacun de ces groupes donne une idée de
leur importance actuelle. Les Arthropodes sont actuellement le groupe qui a
le plus de succès, et ce succès s'explique par celui des Insectes. Certains

4 - Animaux: Structures et Fonctions
BIO 2525 - Hiver 2003
autres embranchements dominent: les Mollusques (moules, escargots,
pieuvre), les Nématodes et les Chordés (dont nous faisons partie, mais
dont plus de la moitié des représentants sont des poissons).
Tableau 1. Nombre d'espèces vivantes dans
les principaux embranchements. (Les 25
embranchements qui ne sont pas inclus
totalisent environ 8,500 espèces
Besoins des animaux
Oxygène
L'oxygène est un élément indispensable à la vie de la plupart des ani-
maux. L'acquisition d'oxygène, qui sert à oxyder des hydrates de carbones
pour produire de l'énergie chimique ou musculaire, est donc une priorité. Le
système respiratoire est l'ensemble des tissus ou organes qui vont permettre
à l'animal de se procurer l'oxygène dont il a besoin.
Les besoins en oxygène d’un organisme sont reliés à sa taille (voir “
Implications écologiques de la taille” à la page 120). En milieu terrestre,
l'oxygène de l'air est disponible en abondance. Dans l'atmosphère, l’oxygène
constitue environ 20% (en masse) de l'air. L'oxygène est relativement peu
soluble dans l'eau (voir “ Solubilité de l’oxygène dans l’eau” à la page 134), et
il est donc relativement difficile pour les animaux aquatiques de se procurer
de grandes quantités d'oxygène. De plus, la solubilité des gaz comme l'oxy-
gène diminue lorsque la température augmente. Le réchauffement d'un
volume d'eau de 5 ºC à 20 ºC lui fera perdre près de la moitié de l'oxygène en
solution. Pour beaucoup d'animaux, dont les besoins en oxygène augmen-
tent avec une augmentation de la température, cela signifie donc que l'acqui-
sition d'oxygène en milieu aquatique lorsqu'il fait chaud est particulièrement
problématique.
Embranchement Nombre d’espèces
Arthropoda 1,200,000
Mollusca 110,000
Nematoda 90,000
Chordata 47,200
Apicomplexa 20,000
Platyhelminthes 15,000
Annelida 15,000
Ciliophora 7,500
Echinodermata 6,000
Sarcomastigophora 4,500
Porifera 4,300

Introduction - 5
Département de biologie, Université d’Ottawa
Eau
Tous les animaux ont besoin d'eau pour survivre. Les réactions biochi-
miques dépendent d’enzymes et de substrats qui doivent être en solution
dans l'eau. De plus, même des processus physiques comme la diffusion de
l'oxygène au travers des membranes requièrent de l'eau. Sans eau, les ani-
maux ne peuvent respirer!
L'eau est également utilisée par les animaux comme solvant pour les
déchets métaboliques. Le gaz carbonique produit par la respiration, et les
déchets azotés produits par le métabolisme des protéines doivent être élimi-
nés. En rejetant l'eau chargée de ces déchets, l'animal peut ainsi maintenir
ses fluides internes propres.
Par définition, l'eau est plus disponible en milieu aquatique qu'en milieu
terrestre. Toutefois, même en milieu marin, l'eau peut être difficile à capter
pour un animal dont les fluides internes sont moins concentrés que l'eau de
mer. En milieu d'eau douce (dulcicole), l'eau peut être trop abondante!
Comme les fluides internes de la plupart des animaux sont plus concentrés
que l'eau douce (ils sont hypertoniques à l’eau douce), l'eau entre continuel-
lement dans l'organisme par diffusion. Cette entrée massive d'eau perturbe
l'équilibre osmotique des cellules, inhibe ou ralentit les réactions biochimi-
ques, et peut même causer l'éclatement des cellules. Les animaux doivent
donc maintenir la concentration en sels et en déchets des fluides internes à
l'intérieur de limites assez strictes: le système osmorégulateur se chargera de
contrôler la pression osmotique et le système excréteur se chargera de puri-
fier les fluides internes en éliminant les déchets métaboliques sous forme
dissoute.
Nourriture
Contrairement aux plantes, les animaux ne produisent pas eux-mêmes
leur nourriture. Ils doivent donc être équipés pour se la procurer dans le
milieu où ils vivent. Les proies capturées devront être digérées et les élé-
ments nutritifs assimilés par l'animal. Le système digestif se chargera de ce
travail. L'élimination des substances qui ne peuvent être digérées est la défé-
cation, distinguée ici de l'excrétion des déchets métaboliques.
Reproduction
L’oxygène, l'eau et la nourriture sont indispensables à la survie des indi-
vidus. Cependant, puisqu'aucun animal n'est immortel, il est indispensable
que les animaux se reproduisent pour assurer la survie de leur espèce.
Fonctions nécessaires à la vie animale
Pour assouvir leurs besoins (voir “ Besoins des animaux” à la page 4), les
animaux doivent respirer, boire, s'alimenter, et se reproduire. Pour bien
accomplir ces fonctions, certaines autres actions sont très souvent nécessai-
res: faire circuler l'oxygène, l'eau et la nourriture vers chacune des cellules,
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
1
/
150
100%