Diaporama

METABOLISME
ENERGETIQUE
CELLULAIRE

INTRODUCTION
On a vu les mécanismes moléculaires des
transports de matière
(diffusion ou protéines transmembranaires,
flux vésiculaire…)

Comment visualiser
les CONVERSIONS d'énergie
de la cellule
les forces qui déterminent
direction et intensité
des déplacements de matière
?

PLAN
I. Structure générale du métabolisme
et rôle des coenzymes
II. Ex. d'anabolisme : la Photosynthèse
III. Ex. de catabolisme : du Glucose

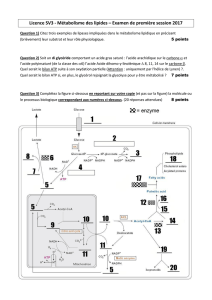
I. Structure générale du métabolisme
II. La photosynthèse : ex. d'anabolisme
III. Ex. de catabolisme d'une molécule
énergétique : le glucose
A. Première étape : glycolyse anaérobie dans le
cytoplasme
B. Deuxième étape : devenir du pyruvate
On se place dans
une cellule animale.
cf. chez cellules
végétale
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
1
/
56
100%