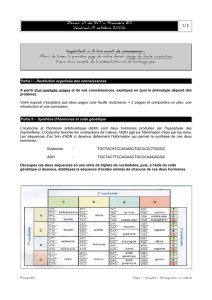
regulation hydrique, ADH


La régulation de l’équilibre hydrique

Bilan journalier de l’eau
L’égalité des gains et des pertes n’est pas un hasard, elle résulte de régulations. Les pertes
sont régulées au niveau rénal et les gains par l’ingestion d’eau liée à la sensation de soif.
boissons
aliments
métabolisme
urine
évaporation
pulmonaire et
cutanée
sueur
excréments
GAINS PERTES

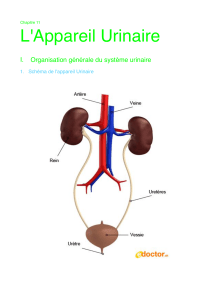
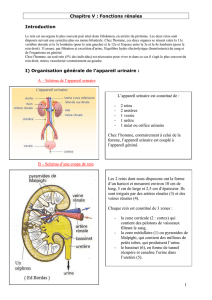
Notions de base sur la structure et le fonctionnement des reins
Chaque rein humain est composé d’environ un million de tubes, les néphrons. Le néphron
constitue l’unité structurale et fonctionnelle du rein.
vessie
urètre
uretère
rein glande surrénale
veine cave inf.aorte
artère
rénale hile
veine rénale
bassinet
néphron
pyramide
rénale
cortex
médulla
uretère
artère
rénale
veine
rénale

cortex
médulla
canal
collecteur
anse de
Henlé
glomérule
tube contourné
proximal tube contourné
distal
capsule de
Bowman
les néphrons peuvent atteindre 5 cm de long mais leur diamètre maximal n’est que de 50 à
60 µm car leur paroi est constituée d’une seule couche de cellules.
canal collecteur en
coupe transversale
lumière
tubulaire
Notions de base sur la structure et le fonctionnement des reins
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
1
/
33
100%