Document

PACES 2012 : UE1 Biochimie
ED 3 :
Acides Aminés
Peptides
Protéines
Enzymologie
Coenzymes
Hémoglobine
Structure glucides et lipides

Question 1
Soit le peptide : Met – Gly – Cys– Lys– Asp –Phe – Ser
A : il comporte un seul acide aminé soufré
B : il est clivé par la trypsine
C : il comporte 2 acides aminés essentiels
D : il absorbe dans l’UV
E : La réaction de Sanger sur ce peptide permet d’identifier la
méthionine.

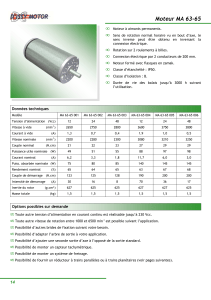
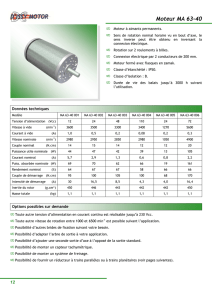
COOH
NH2
CH-
CH2
CH2
S
CH3
Méthionine CH2
SH
COOH
NH2
CH-
Cystéine
A : il comporte un seul acide aminé soufré
Question 1
Soit le peptide : Met – Gly – Cys– Lys– Asp –Phe – Ser
2 AA soufrés : Méthionine (Met) et Cystéine (Cys)

B : il est clivé par la trypsine
La trypsine hydrolyse la liaison peptidique (côté carboxylique) de :
Arg et Lys, 2 AA basiques
Met – Gly – Cys– Lys– Asp –Phe – Ser
Question 1
Soit le peptide : Met – Gly – Cys– Lys– Asp –Phe – Ser
Lysine
(Lys) Arginine
(Arg)

C : Il comporte 2 acides aminés essentiels
Met – Gly – Cys – Lys– Asp –Phe – Ser
Les AA essentiels doivent être apportés par
l’alimentation :
Isoleucine, leucine, lysine, méthionine, phénylalanine,
thréonine, tryptophane, valine
Histidine (grossesse et croissance)
3 acides aminés essentiels
Question 1
Soit le peptide : Met – Gly – Cys– Lys– Asp –Phe – Ser
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
1
/
80
100%