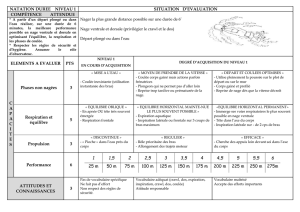
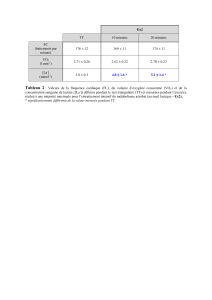
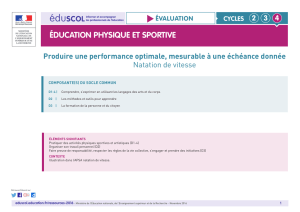
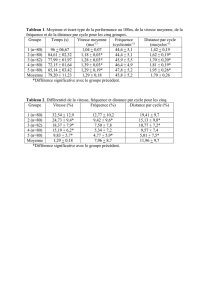
Contribution à l`étude de la sollicitation du métabolisme aérobie au

THÈSE
Pour l'obtention du grade de
DOCTEUR DE L'UNIVERSITÉ DE POITIERS
Faculté des sciences du sport
Unité de recherche Mobilité, Vieillissement, Exercice (Poitiers)
(Diplôme National - Arrêté du 7 août 2006)
École doctorale : Biologie-santé - Bio-santé
Secteur de recherche : Sciences et techniques des activités physiques et sportives
Présentée par :
Chadi Jalab
Contribution à l'étude de la sollicitation du métabolisme
aérobie au cours d'un 100 m nage libre
Directeur(s) de Thèse :
Laurent Bosquet, Olivier Bernard
Soutenue le 28 septembre 2012 devant le jury
Jury :
Président Antoine Gauthier Professeur des Universités, Université de Caen
Rapporteur Antoine Gauthier Professeur des Universités, Université de Caen
Rapporteur Fabrice Prieur Maître de conférences, Université d'Orléans
Membre Laurent Bosquet Professeur des Universités, Université de Poitiers
Membre Olivier Bernard Maître de conférences, Université de Poitiers
Pour citer cette thèse :
Chadi Jalab. Contribution à l'étude de la sollicitation du métabolisme aérobie au cours d'un 100 m nage libre [En
ligne]. Thèse Sciences et techniques des activités physiques et sportives. Poitiers : Université de Poitiers, 2012.
Disponible sur Internet <http://theses.univ-poitiers.fr>

1
THESE
Pour l’obtention du Grade de
DOCTEUR DE L’UNIVERSITE DE POITIERS
Ecole Doctorale : Ecole docotorale BioSanté n°524
Secteur de Recherche :
Sciences et Techniques des Activités Physiques et Sportive (STAPS, 74
ème
section)
Présentée par :
Chadi JALAB
************************
CONTRIBUTION A L’ETUDE DE LA SOLLICITATION DU METABOLISME
AEROBIE, AU COURS D’UN 100 m NAGE LIBRE.
************************
Directeur de Thèse :
Laurent BOSQUET (PU)
Olivier BERNARD (MCU)
************************
Soutenue le 28 septembre 2012
devant la Commission d’Examen
************************
JURY
Antoine GAUTHIER Professeur, Université de Caen Président
Antoine GAUTHIER Professeur, Université de Caen Rapporteur
Fabrice PRIEUR Maître de Conférences (HDR), Université d’Orléans Rapporteur
Laurent BOSQUET Professeur, Université de Poitiers Directeur de thèse
Olivier BERNARD Maître de Conférences, Université de Poitiers co-directeur de thèse

2
REMERCIEMENTS
Au terme de ce modeste travail, je tiens à remercier chaleureusement mon professeur
M. Olivier BERNARD pour la confiance qu’il m’a accordée et pour son accueil amical, ainsi
que ses précieux conseils. Je voudrais lui adresser également mes meilleurs remerciements du
fond du cœur pour sa patience durant ma thèse et pour m’avoir fait bénéficier de ses
connaissances en physiologie du sport. Cette direction a été riche d’enseignements. Il a vu et
vécu toutes les étapes de cette thèse et nous avons finalement atteint notre objectif. Je vous
remercie d’avoir été présent et patient dans les périodes difficiles. Trouvez ici toute ma
reconnaissance et profonde sympathie.
Je remercie tout particulièrement le Professeur Laurent BOSQUET de m’avoir
permis de concrétiser cet objectif universitaire, de son soutien et ses de conseils apportés.
Aux membres du laboratoire (MOVE), grand merci pour votre accueil, encouragement
et soutien dans de nombreuses circonstances.
Le plus grand des mercis aux nageurs, nageuses et aux entraîneurs du stade poitevin
qui se sont prêtés de bonne grâce à mon étude, qui m’ont fait confiance et ont souffert pour
parfois peu de résultats immédiats, conscients que c’est ainsi que la recherche évolue et que
les performances viendront.
À mes parents, rien ne revêt plus d’importance à mes yeux que de vous remercier. J’ai
su apprécier tous vos sacrifices. Merci d’avoir participé même de loin à la réalisation de ce
travail, vous avez été mes meilleurs professeurs, vous m’avez inculqué des vertus et des
valeurs bien plus importantes que toutes les connaissances que j’ai acquises par ailleurs. Vous
m’avez donné toute votre confiance et votre amour. Ce Doctorat et tous les diplômes qui ont
précédé sont aussi un peu les vôtres.
Ma gratitude et mes remerciements vont enfin à mes amis syriens et français qui ont
participé même de loin à la réalisation de ce travail.
Merci à tous

3

4
Résumé :
Le modèle bioénergétique de P.E. di PRAMPERO permet d’expliquer la performance dans
les disciplines de locomotion (course à pied, cyclisme, natation, ...) à partir d’une puissance
métabolique et d’un indice de la dépense d’énergie par unité de distance parcourue.
Néanmoins, pour les épreuves de natation de haute intensité telles que le 100m crawl, le
contexte environnemental rend l’estimation de la puissance métabolique d’origine aérobie
techniquement difficile. L’objectif de cette thèse est alors de proposer une nouvelle approche
des réponses aérobies d’un 100-m nage libre, réalisé dans des conditions les plus proches
possible d’une compétition.
Le travail expérimental a porté sur des nageurs entraînés. Dans un bassin de 25-m, la
procédure propose que les nageurs réalisent une performance sur 100-m nage libre à allure de
compétition et trois épreuves (25m, 50m, 75m) réitérant les mêmes allures intermédiaires que
celles du 100-m. VO
2
est mesuré avant et juste après chaque épreuve, pendant 1min, par rétro-
extrapolation. Les valeurs de VO
2
mesurées à la fin des épreuves de 25m, 50m, 75m et 100m
permettent de reconstruire la cinétique de VO
2
d’une épreuve de 100-m.
Les résultats contrastent avec les estimations des cinétiques de VO
2
publiées précédemment :
VO
2
augmente plus vite dans la première moitié de l’épreuve (à 50m, VO
2
≈ 94%VO
2max
),
atteint 100 %VO
2max
au 75
ème
mètre de l’épreuve puis chute de 7% sur le dernier 25-m. Une
estimation de la contribution relative du métabolisme aérobie montre également des valeurs
plus importantes que celles publiées jusqu’à ce jour, du fait de l’atteinte précoce de VO
2max
en
cours d’épreuve.
Mots clés :
Épreuve sur 100 m nage libre, compétition, cinétique de VO
2
, consommation maximale
d’oxygène, contribution bio-énergétique, méthode de mesure de VO
2
.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
1
/
110
100%