Comparaisons d`abondance de grande faune chassée en

Comparaisons d'abondance de grande faune chassée en Guyane
Française. Résultats préliminaires.
Cécile Richard-Hansen et Benoît de Thoisy*
*Association Kwata
Contexte de l’étude
Dans le cadre du programme d'étude global mené par l'ONCFS et divers partenaires en Guyane
française sur la chasse et son impact sur les populations animales, un réseau de sites plus ou moins
soumis à pression de chasse est prospecté pour quantifier et comparer les abondances respectives des
principales espèces rencontrées.
Méthodes
La méthode utilisée de manière homogène dans tous les sites guyanais est la méthode des line
transects, méthode uniformément adoptée dans la plupart des études en forêt amazonienne (Peres,
1999; Peres & Dolman, 2000).
Tous les sites sont caractérisés par la présence de forêt tropicale sempervirente continue, primaire
ou secondarisée à la suite d'exploitation forestière contrôlée. Les conditions d'observabilité des
animaux sont considérées comme équivalentes dans les différents sites. La pression de chasse est
nulle à très faible dans 6 des zones prospectées : les réserves naturelles des Nouragues et de la
Trinité, les sites très isolés, accessible uniquement par héliportage de Pic Matécho et Piton Baron, et
les zones plus côtières mais à l'écart de voies d'accès comme la zone de Counami Témoin (Counami
T) et Régina Témoin (Régina T). La chasse est présente et régulière dans les autres sites, avec une
intensité variable mais non quantifiable exactement.
Un dispositif similaire composé d'un camp de base (pour les sites isolés) et de plusieurs layons de 3
à 4 km de long, échantillonnant les divers milieux présents a été mis en place dans chaque site. Les
layons sont ensuite parcourus par des observateurs entraînés se déplaçant à une vitesse inférieure à
1 km/h, entre 7h et 11h le matin et 15h –18h l'après midi, de manière répétée jusqu'à cumuler plus
de 100 km / site.
Les résultats sont ensuite traités en termes d'indices kilométriques d'abondance dans un premier
temps (nombre de rencontres par 10 km de transect). Les distances perpendiculaires d'observation
ont également été relevées, permettant dans un second temps d'aborder des calculs de densités, par
la méthode de distance sampling.
Les sites dont les résultats sont présentés dans cette étude ont été échantillonnés, soit par l'ONCFS,
soit par l'association Kwata, en particulier dans le cadre du programme Silvolab "La chasse en
Guyane : vers une gestion durable", réalisé suite à un appel d'offre du ministère de l'Aménagement
du territoire et de l'Environnement (actuel ministère de l’Ecologie et du Développement durable).
Certains sites étaient étudiés dans le cadre d'études pilotes (Counami) ou d'études d'impact (Régina
Saint Georges) menées par l'ONF. Les résultats d'études antérieures sur la Montagne de Kaw
(Julliot, 1998) et de Petit Saut (données pers.) sont présentés également à titre comparatif.
1

Résultats
Comparaison zones chassées-zones non chassées
Les plus grandes espèces étant à la fois les plus recherchées par les chasseurs, et souvent les plus
vulnérables de par leur taux reproducteur généralement plus faible, nous avons considéré dans
l’ensemble des zones échantillonnées les totaux de grandes et petites espèces pour chaque groupe
d’espèces(fig. 1)
0
1
2
3
4
5
6
Matécho
Piton
Baron
Trinité
Nouragues
Régina T
Counami
T
Saül
Counami
chassé
Mont.
Kaw
St Georges
Macouria
(Balata)
Macouria
(Matiti)
Petit Saut
petits primates grands primates
Peu chassé Chassé
0
1
2
3
4
5
6
7
Matécho
Piton
Baron
Trinité
Nouragues
Régina T
Counami T
Saül
Counami
chassé
Mont.
Kaw
St Georges
Macouria
(Balata)
Macouria
(Matiti)
Petit Saut
petits terrestres grands terrestres
Peu chassé Chassé
0
1
2
3
4
5
Matécho
Piton Baron
Trinité
Nouragues
Régina T
Counami T
Saül
Counami chassé
Mont. Kaw
St Georges
Macouria (Balata)
Macouria (Matiti)
Petit Saut
petits oiseaux grands oiseaux
Peu chassé Chassé
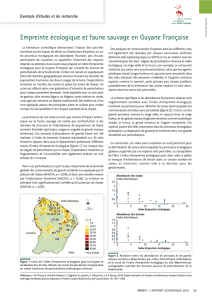
Figure 1 : Abondances relatives des grandes et petites espèces par grands groupes écologiques dans
les différents sites prospectés IKA : nombre de groupes/10km. Les grands primates regroupent les atèles,
hurleurs et capucins, les petits primates les tamarins, saïmiris et sakis. Les petits terrestres regroupent les acouchis et
agoutis, les grands terrestres les pécaris, les daguets et les tapirs. Les petits oiseaux rassemblent les tinamous et les
toclos et les gros oiseaux les hoccos, agamis, et marails.
2

Les abondances (IKA) de grands primates sont toujours plus élevées dans tous les sites non chassés.
Les petits primates, par contre, sont très peu présents à Piton Baron et à Matécho, zones
pratiquement non touchées par l'homme, et beaucoup plus abondants dans certaines zones chassées,
particulièrement la zone côtière de Balata.
La proportion relative des espèces "chassables / non chassables" dans les zones prospectées peut
être un indicateur de la pression de chasse subie, complémentaire de l’évaluation des abondances
sensu stricto. La modification de l’équilibre et de la structure des peuplements de mammifères a
déjà été évoquée dans plusieurs études portant sur l’impact de la chasse en Amazonie (Peres, 1997 ;
Peres & Dolman, 2000). En Amazonie de l’Ouest, une étude comparative (Peres, 1990) avait noté
que la proportion de singes de plus de 4 kg dans la communauté globale de primates passait de 16,4
% en zone chassée à 64 % en zone non chassée. De manière comparable, le long de l’axe Régina -
St Georges, en Guyane, la proportion des espèces "chassables" c’est-à-dire de grande taille, passe
de 90% dans la zone témoin à 44% dans la zone la plus chassée (de Thoisy, 2000). A Saül, la
proportion de ces mêmes espèces est de l’ordre de 63 % de la population de primates, et au Pic
Matécho elle atteint également 90 %. Cet indice doit cependant être pris avec prudence, puisque à
Petit Saut, zone très chassée, la proportion des plus grandes espèces de singes représente tout de
même 64 % du peuplement de primates, malgré des abondances très faibles (0,92 groupes / 10 km
contre 2,2 à Saül, par exemple qui présente un rapport grandes espèces/total équivalent ).
En ce qui concerne les animaux terrestres, l’effet de la chasse est moins évident au premier abord.
Les grands animaux terrestres continuent d’être présents dans certains sites régulièrement chassés,
en abondances souvent supérieures à d’autres zones non chassées comme Piton Baron ou Matécho.
Ces deux zones se révèlent d’ailleurs assez pauvres en petits terrestres également, qui sont souvent
beaucoup plus abondants dans les zones chassées (Saül, Kaw, Counami, St Georges)
De même, les abondances de grands et petits oiseaux varient en fonction des sites, mais pas
forcément de la pression de chasse. Les grands oiseaux se trouvent en abondance réduite à St
Georges et Petit Saut, mais ils maintiennent une population moyenne sur Macouria, voire forte sur
la montagne de Kaw.
Ces disparités d’impact de la chasse sur les différents types d’animaux soulignent bien que la
pression de chasse ne doit pas être la seule raison invoquée pour expliquer des abondances réduites
de certaines espèces dans certaines zones. La quasi-absence de pécaris (Tayassu tajacu et Tayassu
pecari)dans les zones pratiquement intactes de Pic Matécho et Piton Baron, alors qu’ils sont
présents sur le site proche de Saül, chassé depuis de nombreuses années, est un bon exemple qui
incite à rechercher ailleurs les causes sous jacentes responsables de la composition des peuplements
de grands mammifères. De la même manière, les agoutis (Dasyprocta agouti), pourtant chassés,
sont souvent beaucoup plus abondants dans les zones chassées que dans les zones témoins intactes.
Ces premières données comparatives à une échelle territoriale soulèvent donc la question cruciale
du rôle de la structure du milieu sur les abondances des diverses espèces animales. Si la
présence/absence de certaines espèces particulièrement sensibles (singes atèles, Ateles paniscus et
hoccos Crax alector) semble effectivement pouvoir être un indicateur de la pression de chasse, les
populations et peuplements de l’ensemble des espèces dépendent aussi largement de paramètres du
milieu encore difficilement quantifiables.
Variations entre zones non chassées
Cette influence "écologique" sur les densités animales peut se révéler dans un premier temps à
travers l’examen comparatif des abondances relatives dans divers milieux non chassés. En parallèle,
on tentera par la suite de caractériser ces différents milieux selon leur topographie, conditions
pédologiques et géomorpholoqiques et composition floristique, afin de rechercher des paramètres
pertinents sous jacents.
3

Peuplement de primates
En comparant 6 zones soumises à une pression de chasse faible ou nulle, on remarque de manière
préliminaire que les abondances relatives des singes atèles (Ateles paniscus) et hurleurs (Alouatta
seniculus) semblent varier de manière inverse. Les abondances de singes atèles sont effectivement
plus fortes que celles des hurleurs aux Pic Matécho et au Piton Baron, alors que dans les zones
« témoin » de Régina et Counami, ce sont celles des singes hurleurs qui sont plus importantes. La
Trinité et les Nouragues se situent à l’intermédiaire avec des abondances équivalentes pour ces deux
espèces.
Les zones de fort reliefs (Matécho, Piton Baron)semblent particulièrement favorables aux atèles,
alors que les hurleurs prédomineraient dans les zones plus basses et/ou côtières comme Régina,
Counami, et Trinité (Les comptages dans la Réserve nationale de la Trinité ont été réalisés dans une
zone plus proche de fleuves et grandes criques que de l’inselberg). Il faut cependant noter que le
facteur chasse peut quand même avoir une influence sur ce résultat. En effet ces trois dernières
zones ne sont pas chassées régulièrement, ou actuellement, mais sont plus accessibles, et
relativement plus proches géographiquement de zones chassées que les deux premières. Les atèles
étant très sensibles au moindre impact de la chasse, et mettant de très longues années à retrouver un
niveau de population normal après une chute d’effectif (Roosmalen, 1985), il est possible que ces
zones dites "témoin" reflètent tout de même un certain impact de la chasse, passé ou "périphérique"
(Thiollay, 2002). Ayant un taux reproducteur sensiblement plus élevé, les hurleurs sont moins
sensibles à une pression de chasse très faible.
0
1
2
3
4
P. Baron
Matécho
Nouragues
Trinité
Régina T
Counami
T
Alouatta seniculus Ateles paniscus
Figure 2 : Abondances comparées des singes atèles
(Ateles paniscus) et hurleurs (Alouatta seniculus) dans
des sites soumis à pression de chasse très faible à nulle.
Indice Kilométrique en nombre de groupes/10 km
Animaux terrestres
Parmi les terrestres, par contre, les variations sont globalement parallèles pour les deux espèces de
rongeurs d'une part, et pour trois espèces d’ongulés d'autre part. Si ces variations sont en relation
avec des variations de milieu écologique, il semble que les espèces réagissent de manière semblable
aux paramètres environnementaux. La zone de l'Est Guyanais (Régina) semble particulièrement
favorable aux grands animaux terrestres comme les ongulés, alors que les zones à relief marqué
(Matécho, Piton Baron) sont comparativement plus pauvres.
4

Rapport terrestres / primates
Parmi les zones non chassées, on constate que les zones très favorables aux grands primates, et en
particulier aux atèles, sont celles qui ont également de très faibles abondances de grands terrestres,
et particulièrement de pécaris. Là encore, un effet du relief important pourrait être invoqué. On
retrouve ainsi l’indice d’abondance de pécaris le plus élevé (0,88 groupes /10 km) à Saül, pourtant
régulièrement chassé, et relativement proche de Matécho où très peu de pécaris des deux espèces
ont été observés. Il est d’ailleurs à noter que toutes les observations de pécaris à collier de Matécho
ont été réalisées dans les zones échantillonnées les plus basses, au pied du relief proprement dit,
dans une zone de bas fond. Dans la zone de Régina-témoin, au relief collinaire mais moins accusé,
on retrouve également un indice assez élevé de 0,76 groupes/ 10 km .
Cependant, on ne peut réellement conclure encore sur cet effet, tant que plusieurs zones de basse
altitude (forêts en bordures de fleuves, flat), mais non chassées n’ont pas été étudiées. Ce biais
provient de la difficulté d’accès en forêt guyanaise de sites suffisamment éloignés pour ne pas être
touchés par la chasse. Le principal moyen restant l’hélicoptère, des sites autres que les "inselbergs",
zones de reliefs émergents dont le sommet est souvent constitué de roches granitiques affleurantes
permettant un poser en hélicoptère, sont difficiles à trouver.
Ce résultat préliminaire, ainsi que celui des abondances comparées des deux principales espèces de
grands primates, nous amène à considérer comme prioritaire l’échantillonnage de telles zones pour
la suite du programme. Plusieurs sites sont pour l’instant envisagés: la région de l’Armontabo, à
l’Est, la grande pénéplaine du Sud-Ouest, dans lesquelles des "savanes roches" dégagées
permettraient un poser dans une zone non montagneuse, et la zone de l'Arataye, autre secteur de la
réserve des Nouragues situé en bordure de fleuve. Par ailleurs des variations saisonnières dans
l'abondance ou la détectabilité des animaux sont encore à évaluer et à prendre en compte pour
appréhender correctement les facteurs de variation des abondances de faune.
0
1
2
3
4
5
Trinité
P. Baron
Matécho
Nouragues
Counami T
Régina T
Myoprocta acouchi Dasyprocta agouti
0
0,2
0,4
0,6
0,8
Trinité
P. Baron
Matécho
Nouragues
Counami
T
Régina T
Tayassu tajacu Mazama americana
Mazama gouzoubira
Figure 3. Variations des abondances d’espèces terrestres dans divers milieux non chassés (IKA nombres de
rencontre /10 km)
Remerciements
Nous remercions les partenaires et financeurs de ces programmes, et particulièrement la Mission
pour la Création du Parc de la Guyane, la DIREN Guyane, l'ONF et tous les participants à la
collecte des données sur le terrain, tout particulièrement Philippe Gaucher, Isabelle Nolibos et Colin
Niel.
5
 6
6
1
/
6
100%