Enzymologie II La réaction enzymatique La plupart des réactions

Enzymologie II
La réaction enzymatique
La plupart des réactions chimiques survenant dans les cellules se font à des vitesses très
lentes.
Pour observer ces vitesses, il faut des substances qui accélèrent les vitesses de réaction.
Ces substances sont des protéines qui catalysent les réactions, ce sont des enzymes
Quelques définitions :
- Enzyme = catalyseur biologique.
- Catalyseur= substance qui accélère la vitesse d’une réaction mais qui ne
modifie pas l’équilibre de cette réaction
- Catalyseur : il est retrouvé dans le même état à la fin de la réaction, il
n’intervient pas dans la réaction.
- Enzyme= Catalyseur de structure largement protéique. Fixe le substrat avec
une grande affinité et une grande spécificité.
Classification :
- Cellule = environ 4000 enzymes
- Six classes d’enzymes (classifié selon leurs fonctions)
o Oxydoréductases
o Transférases (transferts de groupe chimiques, méthyltranférases
transfert de groupement méthyl … Transporte des groupement
phosphoriques : kinases)
o Hydrolases (Coupures enzymatique : hydrolyses)
o Lyases (Synthétases) capables de liées des groupements sur d’autres
molécules
o Isomérases, réalisent réaction d’isomérisation (modifient places de
différentes fonctions sans modifier … de la molécule
o Ligases (Synthétases) réalise la jonction de deux molécules et qui
nécessite d’apporter de l’énergie généralement car l’action de ligation
est couplée à une hydrolyse de l’ATP
Nomenclature :
- Nom commun
o Trypsine, papaine, pepsine
- Nom de substrat + « ase »
o Uréase, DNAse, arginase
- Nom de la réaction associé au nom du substrat
o Glucose 6-Phosphate déshydrogénase
La vitesse de réaction
La vitesse de réaction ne modifie pas l’équilibre
Keq= (Peq)/ (Seq)=k (Sp)/k (PS)
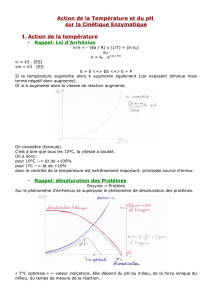
Equation d’Arrhenius

Ln (kT1/kT2)= - (EA/RT)*(1/T1 – 1/T2)
EA= énergie interne
Une énergie d’activation élevée correspond à une très forte augmentation de
température.
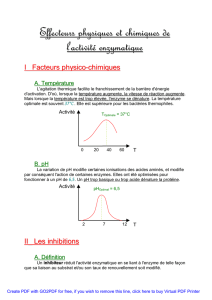
En biochimie, il est très fréquent de mesurer les vitesses de réaction entre 20°C et 40°C,
température ou les enzymes sont actifs.
i (S) = 1 mole
L’énergie cinétique dont sont animées les molécules fournit cette énergie d’activation
Théorie de l’état de transition
S S=/= P
Ν=kT/h dP /dT = (kT/h)(S=/=)
K=/== (S=/=)/(S)= e- (ΔG=/=) /RT
V= fréquence de transformation
K= constante de Boltzmann
H= constance de Planck
On peut interpréter la fréquence à laquelle S donne P
Si équilibre : constante d’équlibre relié à l’énergie d’activation
L’état d’équilibre S activé est démontrer dans bon nombre de cas mais on ne peut pas
l’observer, c’est un état transitoire inobservable, on passe de S à P si énergie suffisante
(S)
le et plus long
S
S
EA= ΔG’

zyme
rapidement
Schéma 1
Soit
Comparons les vitesses des réactions
Pour la réaction non catalysée :
Pour la réaction catalysée :
Soit
enzymatique par rapport à une
Le site actif
-conformation me préfère

Conséquences
mais celui qui peut le faire lors de son état de transition.
Ils se fixent dans une région spécifique : le site actif.
La plupart des enzymes sont très sélectifs dans leur substrat, ils vont reconnaître la
Caractéristique du site actif :
rési
dans la mesure où les enzymes sont de grosses molécules.
Le site actif est donc une entité 3D formée par des groupes qui peuvent venir de résidus
éloignés dans la structure primaire mais rapprochées par la conformation tertiaire de la
molécule. On peut distinguer ainsi 2 types de résidus impliqués dans le site actifs :
Ceux impliqués dans la fixation du substrat.
Ceux impliqués dans la réaction catalytique à proprement dit.
Le
(zinc manganèse cuivre) qui viennent faire des réactions de chélation entre des résidus
afin de rigidifier les sites actifs
Les liaison chimiques qui lient le substrat à
des liaison hydrogènes ou des liaisons ioniques. Dans certains cas il peut y avoir des
interactions hydrophobes. Le site actif est généralement une caverne ou un cillons, le
fixant il peut modifier la protéine et ceci accroît encore
transition.
les
Exemple : la Chymotrypsine.
pour réaliser cette destruction.
1° étape = fixation du su
Dans un certain nombre de cas, il y a redistribution des liaisons
1
/
4
100%