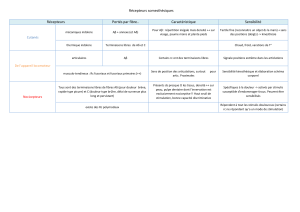
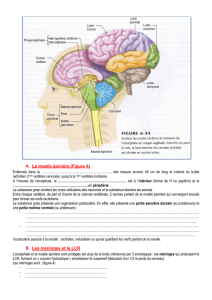
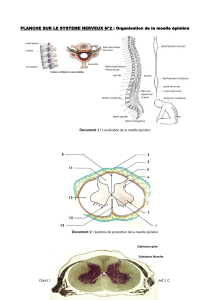
Données anatomiques sur l`organisation des systèmes à monoamines

MINI-SYNTHÈSE
onnées anatomiques sur l'organisation
des systèmes monoamines : un exemple
de communication interneuronale sans synapse ?
m/s n"6, //,juin
(70

1.

�
2.


Ad
1
/
5
100%