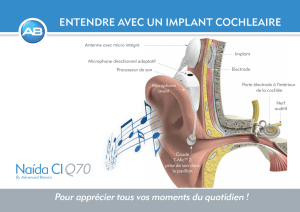
Etude morphologique et fonctionnelle des projections

1
Institut de Physiologie
Département de Médecine
Université de Fribourg (Suisse)
Etude morphologique et fonctionnelle des projections
auditives et non-auditives au noyau cochléaire chez le
cochon d'Inde.
THESE
présentée à la Faculté des Sciences de l'Université de Fribourg (Suisse) pour l'obtention du
grade de Doctor rerum naturalium
Anne-Valérie MONNEY-JACOMME
France

2
Acceptée par la Faculté des Sciences de l'Université de Fribourg (Suisse) sur la proposition de
Raymond Romand, Jean-Pierre Gabriel et Alexandre Babalian.
Fribourg, le ...................................................
Le Directeur de thèse : Le Doyen :

3
Thèse n° 1452
Imprimerie de Miséricorde
2004

4
Acceptée par la Faculté des Sciences de l'Université de Fribourg (Suisse) sur la proposition de
Raymond Romand, Jean-Pierre Gabriel et Alexandre Babalian (experts) et Prof. Marco Celio,
(Président du jury).
Fribourg, le ...................................................
Le Directeur de thèse : Le Doyen :
Prof. Eric M. Rouiller Prof. Dionys Baeriswyl

5
Table des matières
Résumé p. 8
Summary p. 9
Remerciements p. 10
Liste des abréviations p. 11
1. Introduction p. 12
1.1 L'organe récepteur auditif
1.1.1 Structure du système récepteur
1.1.2 Physiologie de l'organe récepteur
1.2 Les voies auditives
1.2.1 Le noyau cochléaire
1.2.1.1 Cytoarchitecture
1.2.1.2 Innervation du noyau cochléaire
1.2.1.3 les voies de sorties
1.2.1.4 Types de réponses des neurones du noyau cochléaire
a. le noyau cochléaire ventral
b. le noyau cochléaire dorsal
1.2.1.5 Tonotopie
1.2.1.6 Autres propriétés fonctionnelles du noyau cochléaire
1.2.1.7 Les principaux neurotransmetteurs dans le noyau cochléaire
1.2.2 Le complexe de l'olive supérieure
1.2.3 Les noyaux du latéral lemniscus
1.2.4 Le colliculus inférieur
1.2.5 Le thalamus
1.2.6 Le cortex auditif
1.3 Notre étude p. 41
2. Matériel et méthode p. 44
2.1 Préparation du cerveau
2.1.1 Animaux
2.1.2 Dissection
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
1
/
123
100%