L`induction du mésoderme - iPubli

SYNTHÈSE
1994 ; JO: 854-67
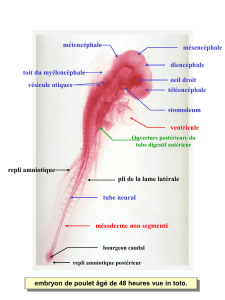
L'induction du mésoderme
à
noggin
Wn
à
goosecoid
à à
à

•

L,
1.
[9].)

32
n

1
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%