II) Etude du cycle de Calvin On peut résumer en 3 étapes le cycle de

II) Etude du cycle de Calvin
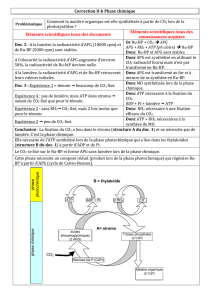
On peut résumer en 3 étapes le cycle de Calvin et Benson
1ere étape : l’incorporation du cO2 dans le ribulose-1,5-bisphosphate
2e étape réduction de l’APG (acide phosphoglycérite) en glycéraldéhyde-3-phosphate
3e étape : régénération
Premièrement le CO2 atmosphérique se fixe à un accepteur en C5 qui est carboxylé (RUBP ou
ribulose-1,5-bisphosphate). Il y a formation d’un intermédiaire en C6 instable, aussitôt hydrolysé en 2
molécules d’acide-3-phosphoglycérique (APG ou 3 phosphoglycérate). C’est le première
intermédiaire stable du cycle de Calvin.
Deuxièmement réduction de l’APG en trioses phosphate. Dans le stroma du chloroplaste, l‘APG est
réduit à la lumière en aldéhyde phosphoglycérique (aldPG ou glyceraldéhyde-3-P). cette réduction se
réalise en 2 étapes, catalysées par la phosphoglycérokinase et pas la glycéraldéhyde-P-
désydrogehase. Au total, la formation de deux molécules de triose consécutives à la fixation d’une
molécule de CO2 nécessite deux molécules d’ATP et deux molécules de NADPH
Troisièmement la régénération du ribulose-1,5-bisphosphate (RUBP)
Le cycle de Calvin permet la régénération du RUBP à partir d’une partie des trioses formés. Ceci va
permettre l’incorporation de nouvelles molécules de CO2. Une partie de l’aldéhyde
phosphoglycérique va être utilisé pour régénérer l’accepteur en C5.
Approche rapide :
1 C5 + 1 CO2 (1 C6) 2 C3,
1 C3 synthèse du saccharose (cytoplasme) ou amidon (chlorolaste).
Pour régénérer le C5 soit à partir d’un C3, possible que si l’on utilise l’équilibre suivant : 5 C3 _ 3 C5,
Le cycle de Calvin se résume donc aux équations suivantes :
3 C5 + 3 CO2 (3 C6) 6 C3 5 C3 + 1 C3 3 C5 + 1 C3
III- Bilan du cycle photosynthétique
On a:
6CO2 +6 ribulose 1,5bisP 12 acide P glycérique
12 acides P glycérique + 12 ATP + 12 NADPH + 12 H+ 12 C3-P +12 ADP +12 NADP++12 Pi
10C3-P +6 ATP 6 rib-1,5-BisP +6 ADP+ 4Pi
2C3-P [hexose] + 2Pi
6CO2 + 18 ATP + 12NADPH + 12 H+ [hexose C6 H 12 O 6 ] + 18 ADP +18Pi +12NADP+
IV- Régulation du cycle de Calvin
V- Synthèse du saccharose
Cette réaction est catalysée par la saccharose-P synthase. Si on isole les chloroplastes et qu’on leur
donne du CO2C14, on n’observe jamais la biosynthese de saccharose. Cependant, si l’on donne du
CO2C14 à des protoplastes, ils fabriquent du saccharose. La biosynthèse du saccharose se fait dans le
cytoplasme. Dans le stroma du chloroplastres les trioses phosphate ald PG et DHAP sont convertis
l’un dans l’autre par isomérisation. Ils sont transportés vers le cytoplasme en échange d’un

phosphate inorganique grâce à des transporteurs spécifiques associés à l’enveloppe du chloroplaste.
Ces transporteurs discriminent préférentiellement le DHAP.
Le DHAP est converti en glucose-1-Phosphate. Ce dernier va être associé à une mocélue très riche en
énergie : l’UDP ou Uridine (ou Uritine ???) triphosphate. Ensuite intervention de la phosphate
phosphatase pour donner du saccharose.
IV) synthèse de l’amidon.
Elle est analogue à la voie de biosynthèse du saccharose. Toutefois a partir du glucose-1-Phosphate,
c’est de l’ATP qui est utilisé et non de l’UDP (ou UTP ??)
La photorespiration
I) Introduction
A) Faits expérimentaux
1) Mesure des intensités de photosynthèse apparente
Avec des plantes en C3, plus on augmente l’intensité lumineuse, plus on observe une augmentation
du CO2 produit (ou de l’oxygène absorbé) en fonction de l’intensité lumineuse. Il existe donc chez les
plantes, une forme particulière de respiration dépendante de la lumière : c’est la photorespiration.
2) Conclusion
Dans le cycle de Calvin, nous avions vu que le Ribulose-5-Phosphate est l’accepteur du CO2 et que
1C5 fixe un CO2 (1C1) pour donner deux C6 : c’est la carboxylation du ribulose-1,5-bisphosphate.
Mais l’enzyme qui intervient (la rubisco) est à la fois carboxylase et oxygénase, elle est donc capable
aussi de fixer l’oxygène. Quand elle agit par sa fonction oxygénase, la rubisco fait réagir le ribulose-
1,5-bisphosphate avec l’oxygène. Cette réaction produit un seul APG (C3) et un C2 (Acide
phosphoglycolique)
II) Origine du CO2 photorespiratoire.
1
/
2
100%