Embryologie du 19 mars 2003

Embryologie du 19 mars 2003.
On s’était quitter au stade phylotypique à la fin de la quatrième semaine du développement.
Au stade phylotypique l’embryon mesure 3 à 4mm. De la fin de S4 à S5 il met en place sa
sensibilité et sa motricité : le développement du système nerveux périphérique.
Mise en place du système nerveux périphérique.
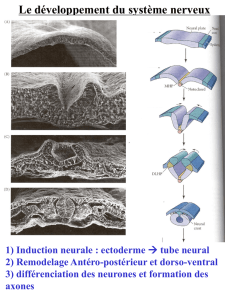
L’ectoderme s’était différencié en plaque neurale puis cette plaque neurale s’est creusé en
gouttière neurale et que les berges de la gouttière neurale sont fait à la jonction sur la ligne
médiane, à peu près au niveau du quatrième somites, et que la fermeture du tube neural qui
s’était faite à la fois en direction craniâl et caudal à partir de point de jonction initiale. La
jonction au niveau du 4eme somites était au J20/21.
De 20 à 24 jours, les parois du tube neural sont simplement constitués d’un épithélium
unistratifié constitué de cellules neuro épithéliale et c’est à partir du J24 qu’on va voir
apparaître une différenciation cellulaire au sein de ce tube neurale. On va analyser
attentivement un schéma : c’est une portion de la paroi du tube neurale avec en bas la lumière
du tube neural et vers le haut la périphérie du tube neural. Initialement, les cellules
neuroépithéliales du tube neural forment un épithélium unistratifié qui occupe toute la largeur
du tube neural. Et le noyau de ces cellules est localisé au périphérie du tube neural. C’est à la
périphérie du tube neural que se situe la membrane basale de cette épithélium et c’est donc le
versant apical des cellules qui est au contact du liquide céphalo rachidien qui est dans la
lumière du tube neural. Ces cellules neuro épithéliales sont dotés d’une capacité de
prolifération intense et lors de la phase S de réplication de l’ADN, les cellules diminuent de
taille en restant amarrées sur leur versant interne qui borde la lumière du tube neural (pole
apical). Ces cellules rondes (en phase S et en mitose) constituent la couche ventriculaire du
tube neural, donc la couche la plus interne, la plus proche de la lumière du tube neural. Cette
prolifération des cellules de la couche ventriculaire va se faire en 3 vagues successives :
1-Une première vague de cellules migre en périphérie de la couche ventriculaire dans la zone
du manteau pour donner les neuroblastes qui se différencieront en neurones de la substance
grise du système nerveux central. Ces neuroblastes émettent des prolongements axonaux vers
la périphérie de la zone du manteau pour former une 3eme couche marginale qui est à
l’origine de la substance blanche du système nerveux central qui ne contient pas de corps
cellulaires neuronaux. Cette description de l’organisation en 3 couches (ventriculaire,
manteau, marginale) est vrai au niveau de la moelle épinière et les choses se compliquent au
niveau du pole céphalique mais nous ne l’aborderons pas.
2-La deuxième vague de cellules prolifératives de la couche ventriculaire se différencie en
glioblastes qui sont les cellules souches gliales (oligodendrocytes et astrocytes).
3-La 3eme vague se différencie en cellules épendymères qui bordent la lumière du tube neural
et qui ont comme fonction de sécréter le liquide céphalorachidien qui remplit la lumière du
tube neural et cette fonction de sécrétion est nécessaire dès lors que le neuropore antérieur et
le neuropore postérieur sont fermés (lorsqu’il était ouvert c’était le liquide amniotique qui
circulait dans le tube neural).
Rapidement on va observer un réarrangement de la zone du manteau au bord de la zone
marginal au niveau toujours de la moelle épinière, pour aboutir à la formation de 4 colonnes.
La lumière du tube neural prend une forme en croix et on identifie deux colonnes ventrales ou
colonnes fondamentales issue de la partie ventrale de la zone du manteau. Ces colonnes
contiennent les motoneurones somatiques qui vont assurer l’innervation des muscles
volontaires striées. Se différencient aussi deux colonnes dorsales qu’on appel aussi colonnes
alaires au niveau desquelles naîtront les neurones d’association entre les terminaisons

axonales des neurones des ganglions rachidiens dorsaux et les motoneurones. Dorsalement
c’est donc sensitive, ventralement motrice.
Un troisième type de colonne se constitue non pas sur l’ensemble de hauteur de la moelle
épinière mais seulement au niveau des métamères thoraciques et des 3 premiers métamères
lombaires (T1 à L3) : il s’agit de la colonne intermédiaux latérale (à la partie la plus dorsal
des colonnes ventrales) qui contient le premier neurone du système sympathique. Cette
colonne se constitue aussi au niveau du 2eme,3eme et 4eme métamères sacrés (S2 à S4) pour
former le premier neurone du système parasympathique sacré.
Le sillon qui sépare les colonnes ventrales et intermédiaux latérales des colonnes dorsales
s’appel le sillon limitant ou sulcus limitans. La lame de tissu neural qui réunit les colonnes
dorsales s’appelle le toit du tube neurale et la lame qui réunit ventralement les colonnes
ventrales s’appelle le plancher du tube neural.
On vient d’étudier la différenciation du tube neural au niveau de la moelle épinière,
maintenant on va étudier la :
Différenciation des cellules de la crête neurale.
Rappel : A la jonction entre les berges de la gouttière neurale et l’ectoblaste, il y avait un
contingent de cellules qui s’individualisent, ce sont les cellules de la crête neurale. Au
moment de la jonction entre les berges de la gouttière neurale, ces cellules de la crete neural
se retrouvent entre tube neural et ectoblaste et ce sont des cellules caractérisées par leur
activité proliférative intense et leur capacité migratoire extrêmement active. Cette migration
va se faire selon 2 voies :
- Une voie superficielle dans le mésenchyme situé juste sous l’ectoderme et ces cellules
des crêtes neurales, lorsqu’elles ont atteintes leur localisation définitive au sein de
l’ensemble de l’épiderme, se différencient en mélanocyte. Ca aide à faire comprendre
le caractère très actif de cette migration. L’embryon est encore de petite taille mais ces
cellules recouvriront toute la surface de l’embryon sauf quand on a une tache
dépigmenté (tache blanche) et là ça signifie qu’il n’y a pas eu migration de ces cellules
des crêtes neurales.
- La deuxième voie de migration est profonde : elle se fait entre le tube neural et les
dérivés des somites et le devenir de ces cellules des crêtes neurales est très différent
selon qu’on se situe au niveau spinal ou craniâl.
le devenir des cellules des crêtes neurales au niveau spinal.
Ces cellules partent de la région dorsale et médial de l’embryon et on va pouvoir décrire 4
arrêts successifs dans cette migration :
Le premier arrêt se fait latéralement au tube neural pour former des ganglions de la racine
dorsale. Les mécanismes de migration et d’arrêt de ces cellules des crêtes neurales au niveau
des ganglions rachidiens dorsaux est bien connu au niveau cellulaire et moléculaire. Au début
les crêtes neurales forment un épithélium et subissent une transformation épithélio
mésenchymateuse avec une diminution de l’expression des molécules d’adhérences cellulaire
N-CAM qui permet leur dissociation les unes des autres. Au niveau des cellules
mésenchymateuses il y a une forte augmentation de la synthèse de la fibronectine qui est une
protéine de la matrice extracellulaire, cette fibronectine est un substrat sur lequel va pouvoir
s’accrocher les cellules des crêtes neurales pour migrer. S’il y a de la fibronectine elles
peuvent s’agripper et suivre le gradient qui les amène sur les bords du tube neural. Arrivées au
bord du tube neural il y a des signaux qui font qu’il y a un arret de synthèse de la fibronectine
dans la matrice extracellulaire, une réexpression des molécules d’adhérences cellulaires N-
CAM et il y a une retransformation mésenchymo épithéliale.

Arrivée à ce stade, les cellules dérivées des crêtes neurales se différencient en neurones de
forme pseudo unipolaires et ces neurones donneront naissance aux fibres nerveuses sensitives
qui conduisent l’influx nerveux depuis les terminaisons nerveuses périphériques vers la
moelle épinière. Deuxièmement en cellules gliales des ganglions de la racine dorsale.
La distribution de ces ganglions de la racine dorsale est segmentaire à l’exception du premier
somite ou métamère cervicale et des 2eme et 3eme métamères coccygiens. On aboutit ainsi à
la formation de 7 pairs de ganglions cervicaux, 12 pairs de ganglions thoracique, 5 pairs de
lombaire, 5 pair de sacrés et une pair de ganglion coccygiens.
Toutes les cellules des crêtes neurales issues de la voie profonde ne s’arrête pas là, il y a en a
qui poursuivent jusqu’au deuxième arrêt.
Le deuxième arrêt se fait latéralement et ventralement au tube neural et ces cellules des crêtes
neurales nées au niveau cervicale, thoracique, lombaire, sacré et coccygiennes (C,T,L,S,C)
forment les neurones et les cellules gliales de la chaîne ganglionnaire sympathique. Les
neurones de cette chaîne ganglionnaire sympathique constitueront les 2eme neurones
sympathiques moteur dont les axones rejoindront le nerf spinal par le rameaux communicant
gris et qui assure le contrôle des fonctions involontaires (rythme cardiaque, sécrétions
glandulaires, mouvements intestinaux) au cours des situations d’alertes, de stress. Alors qu’au
niveau thoracique, lombaire et sacrale il y a formation d’une paire de ganglions au niveau de
chaque métamère, les cellules de la crete neurale cervicale forment simplement 3 paires de
volumineux ganglions sympathiques cervicaux supérieur, moyen et inférieur et les cellules de
la crête neurale coccygienne forment une seule pair de ganglion sympathique coccygiens. Ces
ganglions sympathiques reçoivent les axones des premiers neurones moteurs sympathiques
centraux par les rameaux communicants blancs du nerf spinal provenant des 12 segments
thoracique et des 3 premiers segments lombaires de la colonne intermédio latérale. Le
système sympathique est donc toraco lombaire du point de vue de sa commande centrale. De
T1 à T4 de la colonne intermédio latérale, assure commande sympathique au niveau de la tête,
du cœur et du poumon et de T5 à L3 du tube digestif et de ses dérivés. Ces fibres
préganglionnaires sympathiques qui arrive par le rameaux communicant blanc au ganglion
sympathique ne font pas forcément synapse dans le ganglion sympathique au même niveau :
certaine peuvent se poursuivre cranialement et caudalement avant de faire synapse et
l’ensemble de ces fibres préganglionnaires qui relient les ganglions sympathiques constituent
le tronc sympathique qui assure l’innervation de l’ensemble des ganglions sympathiques. Du
fait du décalage entre origine des premiers neurones sympathiques au niveau donc de la
colonne intermédiaire T1 à L3 et l’origine des deuxièmes neurones à partir des cellules des
crêtes neurales C, T, L, S et C, les ganglions cervicaux, les ganglions L4, L5, sacrés et
coccygiens ne reçoivent pas de rameaux communicants blancs. Tout les neurones
sympathiques périphérique ne sont pas localisés au niveau de la chaîne ganglionnaire
sympathique : un certain nombre de cellules de la crête neurale poursuivent leur migration
jusqu’au niveau de l’aorte dorsale où elles forment les ganglions préaortiques aussi appelés
pré vertébraux : c’est le 3ème arrêt d’où sont issus des fibres sympathiques post ganglionnaires
destinés à l’innervation de l’intestin. Quand on a des douleurs abdominales, elles sont
sourdes, mal localisés car ces cellules des crêtes neurales forment quelques gros amas, pas de
distribution précise. Les cellules des crêtes neurales cervicales formeront une pair de ganglion
à la base du tronc cœliaque. Les cellules des crêtes neurales thoraciques formeront un paquet
de ganglions assez diffus associés à l’artère mésentérique supérieur et les cellules des crêtes
neurales lombaires formeront un paquet de ganglions assez diffus associés à l’artère
mésentérique inférieur. L’innervation préganglionnaire de ces ganglions préaortiques est
assuré par les nerfs splanchniques qui naissent au niveau T5-L3. Les axones à destinée de ces
nerfs splanchniques entrent dans les ganglions de même niveau de la chaîne sympathique et

en sorte immédiatement sans faire synapse pour former les nerfs splanchniques qui se
distribuent aux différents ganglions préaortiques.
La migration la plus longue, celle qui va aboutir au 4eme arrêt, concerne les ganglions
parasympathiques. Ils naissent des cellules des crêtes neurales au niveau des métamères
occipitaux et cervicaux. Ils migrent à travers le mésenchyme/le mésentère de la paroi du tube
digestif depuis l’œsophage jusqu’au rectum où ils forment les ganglions entériques
parasympathiques.
Il y a un deuxième contingent des cellules des crêtes neurales qui naît au niveau des
métamères sacrées et migrent au niveau du mésenchyme qui entoure le rectum et le colon.
L’innervation parasympathique de la partie terminale du colon a donc une double origine :
occipitaux cervical et sacré du point de vue de l’origine des cellules des crêtes neurales. Ces
ganglions parasympathiques entériques sont connectés au système nerveux central par un
premier neurone qui chemine avec la 10eme paire des nerfs crâniens : les nerfs vagues et les
nerfs spinaux sacraux 2, 3 et 4. Le système parasympathique est donc du point de vue de sa
commande par le système nerveux central, craniaux sacrés (alors que pour le système
sympathique sa commande était thoraco lombaire). Ce système parasympathique est actif et
prépondérant dans les fonctions habituelles de maintenance et de digestion et est donc
prédominant pendant les repas et en dehors du stress. De temps en temps les neurones
parasympathiques ne vont pas jusqu’à la paroi de l’intestin du coup des zones de l’intestin se
retrouvent sans ganglion entérique donc ces zones n’ont pas de motricité donc ça cré une
occlusion digestive donc on retire cette partie car ça les empêche de digérer : c’est la maladie
de Hirsprung.
Mise en place du système nerveux périphérique au niveau médullaire.
Les axones moteurs des colonnes ventrales sont les premiers à émerger de la moelle épinière
environ au J30 et ces axones forment la racine ventrale du nerf spinal qui traverse le
sclérotome et tombe sur le myotome et s’y accroche. Puis les axones sympathiques des
neurones des colonnes intermédiaux latérales (T1 à L3) rejoignent la racine ventrale. Quand
les motoneurones passent à proximité des ganglions dorsaux, ceux ci émettent 2
prolongements :
1- Un centripète dirigé vers le tube neural qui fait synapse avec les neurones de la
colonne dorsale
2- Un centrifuge vers la périphérie qui rejoint la racine ventrale et qui va se poursuivre
jusqu’aux terminaisons sensitives définitives.
Remarque : Ce qui migre d’abord c’est le corps cellulaire et quand il est en place y partent des
prolongements axonaux et dendritiques. Comme les cellules sensitives sont des cellules très
polarisés avec des dendrites qui apportent l’influx nerveux vers le corps cellulaire et les
axones qui sortent du corps cellulaire pour aller faire leur synapse, on a toujours tendance à
considérer la morphogenèse comme la mise en place d’abord des terminaisons dendritiques
périphériques en suivant le sens physiologique alors qu’en embryologie c’est le corps
cellulaire qui se met en place puis des prolongements. Ce n’est pas un hasard si ces
prolongements axonaux et dendritiques se mettent à pousser juste au moment où la racine
ventrale passe, c’est qu’en faite celle ci émet un signal qui est reçu par le ganglion rachidien
dorsal, ce signal débute l’émission des prolongements.
L’ensemble ganglion + prolongement dorme la racine dorsale. La réunion de la racine
ventrale et de la racine dorsale forme le nerf spinal ou nerf rachidien.
De T1 à L3, la fibre préganglionnaire issue de la colonne intermédiaux latérale quitte le nerf
spinal par le rameau communicant blanc pour entrer dans le ganglion sympathique
correspondant et comme on l’a déjà dis toute ces fibres préganglionnaires ne font pas relais
dans le ganglion correspondant : certaines rejoignent les ganglions plus antérieur ou

postérieur et ces fibres qui relient l’ensemble des ganglions sympathiques constituent le tronc
sympathique.
Les fibres post ganglionnaires qui naissent de chaque ganglion de la chaîne forme une
branche : le rameau communiquant gris qui rejoint le nerf spinal. Distalement au rameau
communiquant, le nerf spinal contient donc les fibres sensitives, motrices somatiques et
sympathiques post ganglionnaires. Une fois que les axones ont adhérer au myotome, ils
suivent les mouvements des cellules musculaires et les cellules du myotome forment les
muscles de l’épimère qui forment les muscles rachidiens et les muscles de l’hypomère pour
former les muscles du tronc ventro latéral. La première bifurcation du nerf spinal donne les
branches primaires dorsalement à destinée de l’épimère et ventralement à destinée de
l’hypomère (qui donneront les muscles ventraux).
Les fibres motrices post ganglionnaires sympathiques qui innervent les muscles lisses des
vaisseaux sanguins, des glandes sécrétoires, des poils suivent initialement le trajet des axones
moteurs puis quand elles se sont suffisamment approchées des zones dont elles vont assurer
l’innervation, elles vont suivre les trajet vasculaire qui sont déjà constitués.
Les axones sensitifs suivent les voies pré établit par les fibres motrices observe la description
complexe des nerfs en anatomie.
Remarque : L’embryon a du mal à définir une information de position : toutes les connexions
très précises s’établissent à un stade où il y a très peu de liberté donné aux connexions pour
qu’elles s’établissent. C’est les migrations ultérieurs qui vont définir tout ce cablage
complexe.
Au total l’innervation motrice volontaire est segmentaire. L’innervation sensitive est elle aussi
segmentaire avec quelques exceptions puisqu’il n’y a pas de formation des ganglions de la
racine dorsale au niveau de C1 et des 2ème et 3ème segments coccygiens. Donc à peu près
segmentaire avec certaines zones de recouvrement aux limites des dermatomes (=territoires
cutanés innervés par les nerfs sensitifs). L’innervation du SNA par contre est loin d’être
segmentaire.
Niveau céphalique.
Au J28 au stade phylotypique, le cerveau est au stade 3 vésicules :
1-prosencéphale au niveau duquel est constitué un vésicule optique dès ce stade, et ici on
distingue 2 neuromères (mais c’est sujet à discussion).
2-le mésencéphale : 2 neuromères.
3-Rhombencéphale : 9 neuromères dont 8 rhombomères.
Les cellules des crêtes neurales qui naissent en regard de ces vésicules cérébrales ont un
comportement spécifiques : Tout d’abord elles débutent leur migration avant même que le
tube neural ne soit complètement fermé. On décrit ici alors 2 arrêts :
1-La premier arrêt est comme au niveau médullaire, ces cellules des crêtes neurales
fournissent les cellules des ganglions sensitifs des nerfs crâniens.
-Ainsi, les cellules des crêtes neurales qui migrent en regard du 2eme Rhombomère formeront
les ganglions sensitifs de la 5eme pair des nerfs crâniens : le nerf trijumeaux.
-Les cellules des crêtes neurales qui naissent en regard du 4eme rhombomère donneront les
cellules des ganglions sensitifs de la 7eme paire des nerfs crâniens : le nerf facial.
-Au niveau du 6eme rhombomères, les cellules des crêtes neurales vont données les cellules
des ganglions sensitifs de la 9eme paires crânienne : le glosso-pharyngien.
-Enfin, au niveau du 8eme Rhombomères elles vont données des cellules des ganglions
sensitifs de la 10eme paire des nerfs crâniens : nerfs vagues.
2-Le 2eme arrêt de ces cellules des crêtes neurales, c’est pour former les cellules des
ganglions parasympathiques des nerfs crâniens.
 6
6
1
/
6
100%