III. Diversité des chaines TCR - Cours de DCEM1 2010/2011 à Amiens

Immunologie – E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
RECEPTEUR DE L’ANTIGENEE DES LYMPHOCYTES T
- Le récepteur des lymphocytes T est appelé TCR.
- C’est un hétérodimère composé de deux chaines polypeptidiques glycosylées αβ ou γδ liées par
un pont disulfure.
- Chaine chaine possède deux domaines immunoglobuline : un domaine variable (V) et un
domaine constant (C).
- Distribution clonale des TCR : chaque lymphocyte T exprime un type de TCR.
- Le TCR est exclusivement membranaire et possède un seul site de reconnaissance antigénique
(contrairement aux immunoglobulines) : clonotype.
- Dans le sang :
o > 90% TCRαβ.
o <10% TCRγδ.
- Chaque chaine variable comprend :
o Des régions conservées (=Framework).
o Trois boucles hypervariables (HV ou CDR = Complementary Determining Region) qui sont
explosée à la surface de la molécule. Elles sont impliquées dans la reconnaissance
antigénique (Vβ possède une HV4 impliquée dans la reconnaissance des superantigènes).

Immunologie – E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
I. Gène du TCR
- Chaque cahine est le produit du réarrangement (+/-) aléatoire au niveau germinal de segments
géniques : V-(D)-J-C.
- C’est l’une des très rares cellules de l’organisme dont le génome n’est pas complet
(réarrangement au niveau germinal).
- Réarrangements séquentiels DJ puis VDJ.
- Nombreux segments V et J, quelques segments D :
o Réarrangement aléatoire des segments V(D)J qui assure la diversité combinatoire.
o Jonctions V-D et D-J de longueur et de composition variable : N-additions et/ou excision
de nucléotides permettant la diversité jonctionnelle.
- Il n’y a pas de mutation somatique.
- Il n’y a pas de commutation isotypique.
1. Locus α et β
- Pour la chaine α :
o Codée par gènes sur le chromosome 14.
o Codée par le segment Ve tg le segment J qui peuvent se recombiner de manière
aléatoire. Diversité combinatoire environ 60 x 60.
- Pour la chaine β :
o Codée par locus sur chromosome 7.
o 50 gènes V et 2 Clusters avec chacun segment V, J et C.
- Segment Vα t Vβ regroupés en familles (>75% homologies).
2. Locus δ et γ
- La diversité combinatoire de la chaine Cδ est faible car peut ... .
- Particularité du locus delta : inséré dans le locus α.
o Petit cluster D J et C est inséré entre les segments V et les segments J.
o Explique qu’au cours de l’oncogénie des lymphocytes T il y a d’abord un réarrangement δ
a puis un α.
- Locus δ inclut dans le locus α : délétion lors du réarrangement α.
- 8 Vδ dont 5 Vα (communs).
- 2 segments Dδ peuvent être utilisés simultanément (long CDR3).

Immunologie – E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
- Diversité combinatoire réduite.
- Importante diversité jonctionnelle (chaine δ).
3. Mécanismes du réarrangement du TCR
- Réarrangement séquentielle :
o D’abord D sur J : intervention de protéines particulières tels que RAG1, RAG2, DNA-PK,
Artémis, etc. Pathologie : Si RAG non fonctionnel absence de LT et LB.
o N et P addition grâce à la TdT.
o Puis réarrangement aléatoire de V sur segment DJ recombinés. Intervention des mêmes
enzymes que précédemment. Toujours des N et P additions grâce à TdT.
o Il y a alors une transcription.
o Le pré-ARNm subit un épissage.
o L’ARNm sera traduit en protéine.
4. Expression clonale du TCR
- Réarrangement de la chaine β par exclusion allélique :
o Réarrangement séquentiel sur les deux chromosomes (un allèle à la fois, jamais les deux
allèles recombinées en même temps).

Immunologie – E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
- Réarrangement de la chaine α se fait sans exclusion allélique.
o Les deux allèles se réarrangent simultanément.
o Il y a une probabilité que 1/3 des LT expriment deux chaines α (mais une seule chaine β).
Il y a donc deux types de TCR.
o En situation physiologique un seul de ces deux TCR est fonctionnel. Donc malgré
l’expression de deux TCR il reste expression clonale.
o Dans des situation pathologique le deuxième TCR peut devenir fonctionnel, donc les LT a
deux TCR sont des facteurs de risque pour l’apparition d’une auto-immunité.
5. Réarrangement Vα-Jα
- Certaines séquences spécifiques présentent sur le locus permet la formation d’une boucle sur
elle même de la partie excisée.
- Cercle d’excision du LT (appelée TREC) se comporte comme un épisome.
- Une cellule comprenant un TREC :
o La cellule se divise et donc le matériel génétique se réplique sauf le TREC (car épisome).
o Le nombre relatif de TREC diminue au long des divisons cellulaires.
- La quantification des TREC permet d’évaluer les émigrants thymiques récents: mesure de
l’activité du thymus. Il est possible de quantifié les TREC afin d’évaluer la production thymique.
o Quand le thymus fonctionnement bien, il y a de nouveaux LT avec pleins de TREC. Les
vieux LT qui se sont divisés ne possèdent plus beaucoup de TREC.
o Utilité dans le suivi thérapeutique.
II. Répertoire lymphocytaire T
- Le répertoire lymphocytaire T correspond à l’ensemble des TCR présent chez un individus.
- Il est différent d’un individu à l’autre et n’est pas prédictible avec le génome.
- Le répertoire théorique est de l’ordre de 1015 à 1018 (ce qui contribue le plus à la diversité totale
est la diversité jonctionnelle).
- Pourquoi théorique ? car jamais aussi important en effet il y a des contraintes exercées sur les
répertoires :

Immunologie – E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
o Sélection positive et négative. Phénomènes qui ont lieu pendant le développement des
lymphocytes T.
o Contraintes stériques d’associations α+ β. Deux chaines β et α au hasard peuvent ne pas
s’associer par problème de structure.
o Réarrangements non aléatoires : tous les segments peuvent êtres utilisés mais certains
sont utilisés de manières préférentielles.
o Nombre de lymphocytes total : nombre de LT chez l’homme (1012) donc on ne peut pas
avoir plus de 1012 LT différents.
- Le répertoire réel des lymphocytes sanguin est de l’ordre de 2.107.
- Or dans le nombre de LT sanguins est de 5.109, donc dans le sang un clone comprend environ 250
cellules.
III. Diversité des chaines TCR
- Domaine variable des chaines β :
o Segment V + segment D + segment J.
o Les trois boucles hypervariables exposées à la surface et impliquée à la reconnaissance
antigénique (appelées CDR1, CDR2 et CDR3).
- Les CDR1 et CDR2 sont spécifiques d’un segment V. Exemple : Vβ6 on connait sont CDR1 et sont
CDR2.
- Le CDR3 lui inclut segment D + segment J + nucléotides entre VD et entre DJ.
o Variabilité combinatoire et jonctionnelle.
o Spécifique de chaque chaine V.
o Exemple : deux chaines Vβ.2-Dβ1-Jβ1.2 peuvent avoir des CDR3 différents.
- Le CDR est dont la région la plus variable du TCR.
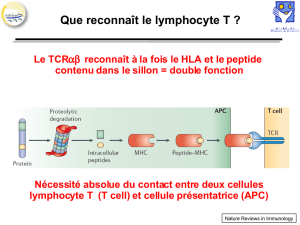
IV. Présentation antigénique
- Les CDR1 et CDR2 des chaines α et β sont principalement en contact avec les sillons α qui bordent
le sillon à peptide du CMH.
- Les CDR sont principalement en contact avec le peptide central.
- Le complexe CMH/peptide polymorphique est en contact aves les régions hypervariables du TCR.
- Logique car :
o Un individu donné a 6 molécules HLA classe I différentes et 6 molécules HLA classe II
différentes (donc assez restreint) s’assemble avec CDR1 et CDR2 de nombres différents
restreint car codé par le génome.
o Le peptide qui varie selon chaque situation pathologique lui s’associe avec la partie la
plus variable du TCR : CDR3.
 6
7
6
7
1
/
7
100%