4eme cours: 10 oct

Le 10/10/05 M. CHAMBOST et T. BOULET
Rappel :
-Les lymphocytes (Ly) T et B sont des cellules dites immunologiquement compétentes. Elles
ont une réponse spécifique de l’antigène (AG).
Les LyB reconnaissent l’AG natif et soluble grâce à leur
immunoglobuline (Ig) de surface.
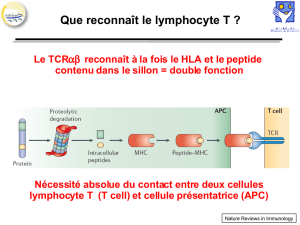
Les LyT reconnaissent l’AG présenté par une cellule présentatrice
de l’AG (CPA) grâce au TCR. L’AG es présenté (sous forme
dégradée) par une molécule de CMH
De classe I pour les T CD8+
De classe II pour les T CD4+
-Dans la moelle osseuse les LyT n’ont pas de TCR.
Dans le thymus il y a :
acquisition du TCR
double sélection (positive et négative)
°Chaque LyT a un TCR unique et original
°La sélection positive sélectionne les LyT ayant un TCR capable d’interagir avec le
CMH de l’individu.
°La sélection négative élimine les LyT ayant un TCR auto-réactif.
°Si l’affinité du TCR pour le CMH est très forte, le LyT reçoit un signal négatif. En
effet, trop d’affinité entraîne une reconnaissance des peptides du soi.
Seuls les LyT ayant un TCR dont l’affinité pour le CMH est moyenne seront
recrutés.
Il faut comprendre que le thymus étant un organe lymphoïde central il est déconnecté de
l’extérieur, il n’y a donc pas d’AG étrangers.
Ici il n’y a présentation que de peptides du soi. C’est pour cela qu’une affinité TCR/CMH trop
forte entraîne la délétion du LyT en question.
Dans les organes lymphoïdes périphériques, au contraire, une interaction TCR/CMH donnera
toujours un signal positif.
Le caractère Adaptatif de la Réponse Immunitaire
1/ Adaptation au soi :
Il existe une double adaptation à l’individu
La sélection positive : Les TCR des LyT sélectionnés sont capables
d’interagir avec les molécules du CMH de l’individu selon une
affinité moyenne.
Remarque : Quand il y aura reconnaissance d’un AG l’affinité peut devenir très forte.

La sélection négative : On dit que ces sélections se font à partir du
« répertoire primaire » des LyT
2/ Adaptation au milieu extérieur :
Quand il existe une affinité entre l’AG et le Ly il y a déclenchement d’un signal positif qui
entraîne :
une phase de prolifération
une phase de différenciation en cellule effectrices et en cellules
mémoires.
Les effecteurs peuvent être des T CD4 ou des T CD8 et ont pour but de tuer les
cellules infectées par un virus, une bactérie...
Remarque : si question sur la réponse cellulaire attention de ne pas oublier les mots-clés :
TCR, spécificité et adaptivité.
3/ Le TCR :
C’est une protéine transmembranaire du LyT dotée d’un domaine constant et d’un domaine
variable.
Le TCR est un hétérodimère qui peut être de la forme / ou plus rarement /.
Il existe donc des domaines dits C, C, C, C, V, V…
Le TCR interagit par ses parties variables avec la molécule du CMH.
En réalité il y a une double interaction :
reconnaissance du peptide par les parties variables
Interaction du TCRec les berges de la gouttière du
CMH (interaction possible grâce à la sélection
positive, cette moyenne affinité)
-On ne peut pas transfuser des LyT.
En effet les molécules du CMH sont tellement variables que la probabilité de trouver deux
individus HLA identiques (mis à part les vrais jumeaux) est quasi-nulle.
Les LyT du donneur seraient incapables de reconnaître les AG présentés par les molécules du
CMH du receveur.
-Un des problèmes dans la greffe de moelle est que le thymus du receveur est vieux et n’est
pas capable d’abriter les nouveaux LyT fabriqués par la moelle (car le thymus régresse à
partir de 6-7 ans)
-Dans le cas du SIDA les patients perdent très vite leurs T CD4 mémoires. Or il faut
beaucoup de temps pour régénérer une mémoire immunitaire (en raison de la régression du
thymus). La reconstruction des cellules perdues est longue et imparfaite.
Le complexe de transduction CD3
Le CD3 est un complexe multimoléculaire associé au TCR.
Il est composé de chaînes (, , , ) qui possèdent des domaines intracytoplasmiques plus ou
moins longs.
Dans ces domaines on trouve les motifs ITAM (Immunoreceptor Thyrosine-based Activation
Motif)

A l’intérieur des ITAM des thyrosines seront phosphorylées par des thyrosine kinases.
C’est la phosphorylation de ces motifs ITAM qui donne le signal d’activation.
Deux types de thyrosine kinases sont impliquées : LCK et FYN
Elles sont situées juste sous la membrane cytoplasmique.
LCK est associée au domaine intracytoplasmique de CD4 et CD8.
L’interaction entre les deux cellules excite LCK (interaction entre LyT et CPA). L’excitation
de LCK est complétée lorsque CD45 interagit avec la CPA.
CD45 est une phosphatase qui clive des phosphates appartenant à FYN et à LCK.
FYN et LCK ainsi activées vont phosphoryler les tyrosines des motifs ITAM.
ZAP 70 qui est collée au complexe est à son tour phosphorylée.
Elle devient alors à son tour une kinase :
-Elle va activer la protéine oncogène RAS qui active la voir des MAP kinases
-Elle active la phospholipase C spécifique des LyT qui active à son tour la voie de
transduction du signal passent par le PiP2 membranaire clivé en DAG + IP3
->DAG active une protéine kinase C qui va libérer NFKB du complexe IKB-
NFKB (I pour Inhibiteur)
NFKB libre va activer tout un tas de gènes (par exemple le gène de l’IL2)
->IP3 va provoquer l’ouverture des canaux calciques.
Passage du Ca du RE dans le cytoplasme
Entrée du Ca extracellulaire dans la cellule
o Augmentation du Ca intracellulaire
Cette augmentation induit toujours un signal pou toute les cellules
Pour les LyT, le signal post-calcique passe par la Calcineurine (associée à la Calmoduline).
Remarque : La Calcineurine est la seule voie post-calcique du LyT alors que dans les autres
cellules il y a souvent plusieurs molécules qui entrent en jeu.
La Calcineurine devient une phosphatase (Calmoduline dépendante) et elle va retirer un
phosphate à la NFAT.
NFAT non phosphorylée peut alors être transloquée dans le noyau et activer des gènes (leurs
promoteurs) qui possèdent un motif NFAT-sensible.
Il existe des drogues Immunosuppressives qui contrecarrent cette voie.
Ex : La Cyclosporine, peptide cyclique produit par un champignon, se lie à une
immunofiline et ce complexe inhibe la Calcineurine.
Remarque : Concernant le complexe de transduction CD3 :
-Tous les signaux ne passent pas par CD3 et le TCR !
-On ne sait pas vraiment comment le signal passe du TCR au CD3.
-A chaque contact entre LyT et CPA il y a interaction avec plusieurs TCR du
LyT.
Tous les TCR sont rassemblés au même point de la cellule ce qui permet une
somation des signaux.
C’est la simultanéité des signaux qui va activer réellement la cellule.
-On ne sait pas qui est vraiment l’acteur principal (LyT ou CPA) dans ce
mécanisme de rassemblement.
Le co-signal donné par la molécule CD 28

CD 28 est une molécule exprimée par la CPA.
Celle-ci interagit avec CD 28 par deux types de molécule :
° B 7
° CTLA 4
Les deux molécules dont antagonistes.
L’interaction entre B 7 et CTLA 4 induit un signal cellulaire qui s’additionne aux signaux
transmis par l’interaction entre CD 3 et le TCR.
L’interaction entre CD 28 et CTLA 4 induit un signal négatif.
La CPA n’exprime au début que la molécule B 7, CTLA 4 n’existant pas.
Une fois que la CPA a interagit avec la LyT, elle exprime alors CTLA 4 ce qui permet une
balance, un équilibre de l’activation cellulaire.
En effet, plus une cellule aura été activée, moins elle sera activatrice.
Remarque : Il existe des drogues CTLA 4 solubles recombinantes.
1
/
4
100%