Chapitre 2 : Plasticité cérébrale

1
Chapitre 2 : Plasticité cérébrale
Définition du concept de plasticité (J.Paillard, 1976) : la plasticité est la
propriété qu’ont les corps déformables de changer leur forme sous l’action de
forces extérieures et de conserver cette déformation lorsque cette force
extérieure a cessé d’agir.
La plasticité est la caractéristique essentielle du système nerveux de tout être
vivant. Le système nerveux se remanie donc souvent, c’est sa propriété
adaptative.
I – La plasticité au cour du développement
Quelques théories se sont penchés sur le développement du système nerveux.
Trois principales ont été posées à la fin du 19ème siècle :
- La théorie du développement par instruction l’idée que le cerveau a
une grande plasticité, qu’il possède de nombreux mécanismes.
d’apprentissage et que cela lui permet de s’adapter à l’environnement.
- La théorie du développement par construction l’idée est que c’est
non pas l’environnement mais au contraire le système nerveux qui
impose son fonctionnement, ses structures.
- La théorie sélectionniste le développement par sélection
1) La théorie sélectionniste du développement cognitif
Le développement consisterait en une sélection au cours des interactions avec
l’environnement de possibilité compatibles avec cet environnement et en une
élimination des autres.
L’interaction avec l’environnement est nécessaire.
Le système est à la fois pré-formé et en évolution incessante.
Dans le développement du cerveau, il faut prendre en compte la complexité de
l’organisation qui va permettre la mise en place de différentes fonctions.
Il va falloir que se mette en place chez les individus un schéma général
d’organisation (on a tous une moelle épinière, un hypothalamus,…), les
connexions entre les différentes structures, ainsi que la grande variabilité
interindividuelle comme par exemple le poids du cerveau (mais n’indique pas
les capacités mentales), la taille et volume des systèmes cérébrales (variant de
plusieurs mm),…
Ce sont les instructions génétiques qui vont être à l’origine de l’élaboration du
système nerveux central et de l’invariance de l’organisation cérébrale d’un
individu à un autre mais le programme génétique ne va pas s’étendre aux détails
de l’organisation du cerveau.

2
Il va falloir introduire la notion d’épigenèse. Les mécanismes épigenétiques sont
des mécanismes qui vont prolonger l’action des gènes, ils représentent un niveau
supplémentaire d’organisation.
Rappel sur le développement du système nerveux
L’analyse au niveau cellulaire du développement du système nerveux central
permet de distinguer 5 phases successives :
- multiplication & différenciation
- migration cellulaire
- croissance axonale et dendritique
- synaptogénèse
- myélinisation
Ces phases vont être responsable des changements au niveau macroscopique.
Au cour du développement, à 20 jours on voit le tube neural, à 40 jours il donne
3 vésicule dans la partie antérieur et en fonction du taux de multiplication des
cellules ces vésicules vont se développer.
Au cour du développement on compte 2 phases :
- première : les facteurs génétiques et endogènes contrôlent le
développement
- deuxième : l’activité dans les réseau va contrôler le développement
Première phase
Elle va concerner la période de multiplication & différenciation et des
migrations.
Une fois que les cellules sont à leur emplacement définitif, elles vont devoir
faire des contacts, donc on a à la fois le développement des dendrites et des
axones, ces derniers vont devoir entrer en connexion avec d’autres neurones.
Du corps cellulaire vont partir des prolongements qui vont faire contact avec un
neurone, situé + ou – prés. Cela est coder génétiquement mais joue aussi
l’environnement.
Le cône de développement (qui part de la cellule) va être sous l’influence de la
cellule cible, c’est-à-dire que la cellule cible sécrète une molécule qui va être
nécessaire à la survie du neurone qui a le cône de croissance. Un des facteurs de
survie important est le NGF qui est là pour bloquer les mécanismes qui
conduisent à la mort neuronale (apoptose).
Ce que l’on va observer au cour du développement est qu’un grand nombre de
neurone vont disparaître (régression neuronale).
Ces molécules ont un autre rôle qui est de stimulé la formation des
prolongements.
Cette mort neuronale a été mis en évidence par rapport au niveau des ganglions
spinaux.

3
Au cour du développement, les ganglions spinaux ont tous le même nombres de
neurones, puis on va observer que certains vont diminuer en taille, ce sont ceux
qui contiennent les neurones qui innervent le dos et le ventre.
Le nombre de neurone n’est pas codé génétiquement.
Au niveau cortical, à 2 ans au niveau du cortex frontal, l’enfant à 2 fois + de
neurone que par rapport à l’adulte, et encore 10% à 10 ans.
La stratégie est donc de créer un surnombre de neurone puis de les réguler via la
mort neuronale.
Il existe 2 zones dans le cerveau où il y a mort et renouvellement des neurones :
bulbes olfactifs et une zone de l’hippocampe.
Deuxième phase
Une fois que le cône de croissance a rencontré la cellule cible et que les cellules
se forment, le processus de développement va continuer. Là on va observer
qu’un grand nombre de synapse vont se former et qu’un grand nombre vont
disparaître.
Ce qui est à l’origine de cette agencement c’est l’activité dans ces nouveaux
réseaux qui vont contrôler son organisation. Cela a d’abord été mis en évidence
au niveau de la synapse entre les neurones moteurs et les muscles, ce qu’on
appelle la tension musculaire.
Chez l’adulte, chaque fibre musculaire ne fait synapse qu’avec un seul neurone
moteur alors qu’au cour du développement il y a 4/5 neurones qui profilent sur
chaque fibre musculaire. Si on diminue l’activité du muscle on maintient la poly
innervation.
L’axone se met en contact avec la fibre musculaire qui va libéré de
l’acétylcholine permettant la création de la synapse. Dans cette phase là on
observe plusieurs terminaisons qui vont venir permettre la contraction
musculaire. Au final cela créer la synapse qu’on voit chez l’adulte bien efficace.
Ainsi on passe d’une phase où les synapses sont fragiles/labiles à une phase où
les synapses sont stabilisés.
A partir de là est apparu que ( ?) faisait l’objet d’une régulation épigenétique,
que le câblage est déterminé génétiquement mais que c’est l’activité qui règle les
détails.
Au niveau cérébral, le bébé naît avec un cerveau d’environ 300 grammes
représentant environ 1/5ème de son poids d’adulte. Sa croissance va être lié à la
poussée des axones et des dendrites. On voit une augmentation très importantes
du nombre de synapse qui va laisser place petit à petit à une diminution de la
densité des synapses qui va se poursuivre jusqu’à l’adolescence et enfin le
développement des gaines de myéline jusqu’à 20 ans.
Il faut savoir qu’il y a un certain nombre de connexion qui vont se mettre en
place et donc disparaître. Par exemple au départ toutes les zones du cortex
envoient des axones vers la moelle épinière, chez l’adulte il n’y a plu que le

4
cortex moteur. Un autre exemple est sur le cortex visuel droit qui projette au
début sur le gauche et réciproquement, chez l’adulte il ne reste plu qu’une petite
bande proche de la ligne médiane où il y a des connexions entre les deux cortex,
la plupart des autres connexions ayant disparues.
Il y a un décalage temporelle dans la maturation des différentes aires corticales.
On le sait depuis longtemps mais se sont les durées qui ont changées avec les
études. Au départ on pensait qu’avec le cortex visuel le maximum de synapse
était vers 3/4 mois, pour le cortes frontal vers 3/4 ans ; aujourd’hui on prolonge
longtemps ces durées.
Les premières aires à arriver à maturation sont les zones sensorielles et les zones
motrices, on observent une réduction de la substance grise (du nombre de
synapse) entre 4 & 8 ans au niveau de ces zones.
Vers 11/13 ans on observe une diminution de la substance grise mais au niveau
pariétal cette fois-ci (vision global d’une représentation, perception de l’espace).
Le cortex pré-frontal (rôle dans les comportements sociaux, la mémoire de
travail, les prises de décision, ) est la zone qui va présenter une maturation à
l’adolescence.
La dernière zone cortical a montré sa maturation est le cortex temporal
(audition, langage, l’hippocampe).
Dans la maladie d’Alzheimer, la dégénération se fait dans le sens inverse de la
maturation (se finit par les zones sensorielles).
Ce n’est pas parce qu’une zone n’a pas fini sa maturation qu’elle n’a pas
d’activité fonctionnelle, on observe à l’IRM de l’activité lors de présentation de
stimuli visuelles par exemple.
Rôle de l’environnement dans le développement des cortex sensoriels
Au niveau des cortex sensoriels c’est l’environnement qui va être responsable de
l’activité dans les réseaux. Les signaux de l’environnement auront une action
importante sur l’agencement des connexions synaptiques que si ils ont lieu
pendant une période particulière qui s’appelle la période critique.
La période critique/sensible est appelé ainsi quand on a une relation importante
entre l’âge et les facultés d’acquisition. D’un point de vue neurobiologique cela
va être la période transitoire pendant laquelle la connectivité est maximale et
redondante, c’est la période pendant laquelle la distribution des connexions va
s’adapter. C’est la période pendant laquelle des expériences, des conditions
anormales, vont avoir des conséquences irréversibles.
Il existe une période critique en ce qui concerne la vision de la profondeur. Le
système sensoriel visuel fini sa maturation vers 11/13 ans.
Le champ visuel peut être divisé en deux hémisphères visuels, le gauche et le
droit. On peut diviser la rétine en deux parties, la rétine temporale et la rétine
nasale. Des stimuli dans présent dans l’hémisphère visuel gauche active la rétine

5
temporale droit et la rétine nasale gauche. Les axones des neurones des rétines
temporales vont se projeter du même côté d’où ils sont situé alors que les rétines
nasales projettent de l’autre côté.
Au niveau du thalamus visuel droit, il va recevoir les informations des 2 yeux
mais d’un seul hémichamp visuel (le gauche) et réciproquement.
Au niveau du thalamus les informations qui proviennent des 2 yeux se
répartissent sur différentes couches, une pour l’œil droit et une pour l’œil
gauche.
Ensuite du thalamus vont être projeter au niveau du cortex visuel primaire du
même côté (thalamus droit => cortex droit). Le cortex est constitué de 6
couches, celle d’entrée des informations est la couche 4. A ce niveau, les
informations provenant des deux yeux ne se mélangent pas, elles arrivent dans
des territoires différents.
Colonne de dominance oculaire : présente au niveau de cette couche 4, elle ne
reçoit que l’information d’un des yeux, on dit qu’elle est monoculaire.
Au niveau de la couche 3, qui est celle qui reçoit les informations de la couche
4, les informations vont se mélanger, il y aura un certain nombre de neurone qui
recevront des informations de neurone qui sont dans des colonnes de dominance
oculaire différentes. Ces neurones répondent donc aux deux yeux et sont ainsi
dit binoculaires.
A la naissance on a des différences par rapport à l’âge adulte, en effet il n’y a
pas de colonne de dominance oculaire (les neurones reçoivent les informations
des 2 yeux, ils sont donc tous binoculaires). Les cellules de la couche 3 sont
toutes binoculaires également.
L’organisation va se mettre en place au cour du développement.
De nombreuses études ont montrées que la mise en place de l’organisation
dépendait de l’expérience visuelle précoce. Des privations ou des anomalies
visuelles précoces ont un effet sur la mise en place du système visuel.
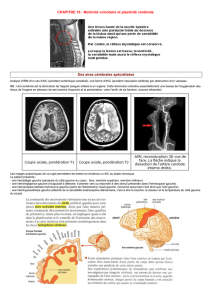
Les effets des privations visuelles
En ce qui concerne le système visuel, on la particulièrement étudié chez le chat
qui est l’animal ayant le système visuel le + proche de l’humain.
Chez le chaton il a été montré que une semaine de fermeture d’un œil induisait
une cécité de l’œil (déficience). Pour observer cet effet là il faut que la privation
monoculaire (fermeture de l’œil) est lieu avant le 4ème mois, la période critique
est donc de la naissance (enfin le 10ème jour quand ils ouvrent les yeux) au 4ème
mois. Au cour de ces mois la sensibilité est maximale au début de la période
critique et puis diminue au fur et à mesure. Si la privation visuelle d’un œil à
lieu au cour de la première semaine, si un reste fermer pendant une semaine, il
va perdre la vision de cet œil et ne va jamais la retrouver (alors que la période
critique se termine au 4ème mois). Cependant si pendant la période critique on
ferme l’autre œil, on observe une restauration de la vision de l’œil qui avait été
 6
7
8
9
10
11
12
13
14
15
6
7
8
9
10
11
12
13
14
15
1
/
15
100%