BIOACOUSTIQUE : FONCTIONNEMENT DE L`OREILLE I) OREILLE

BIOACOUSTIQUE :
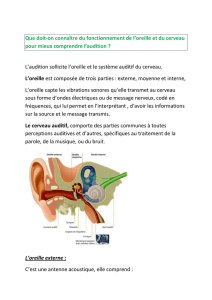
FONCTIONNEMENT DE L’OREILLE
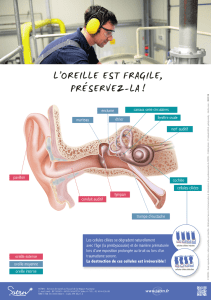
I) OREILLE EXTERNE
La principale fonction de l’oreille externe est la captation et l’orientation des ondes sonores vers l’oreille
moyenne. Toute fois, sa structure offre une sélection préliminaire des fréquences auditives. D’une part elle
amplifie l’intensité des ondes sonores utiles (du langage) ayant une fréquence comprise entre 1 et 5 Khz et
d’autre part elle diminue l’intensité des fréquences nuisibles (10Khz).
Remarque : La perception du son s’accompagne d’une aide à la localisation de la source, par rotation réflexe
et inconsciente de la tête, dans le sens le plus favorable à la pénétration des ondes sonores. L’orientation
auditive (différence de temps interaural) se réalise avec un retard temporel d’environ 2x10-4 secondes.
1,7x10-4 s pour un son qui arrive du coté droit avec un angle de 15° par ex.
Enfin les interférences constructive ou destructive de l’oreille externe sont plus efficaces pour les sons
venant de face que de haut.
• Le pavillon
(lame fibrocartilagineuse faisant
un angle de 30° avec la tête)
• La conque le conduit auditif
externe (C.A.E)
• Chez l'être humain, le conduit auditif
externe a une longueur de 2 à 3 cm et
un diamètre de 0,7 cm.
• L'ensemble de la conque et du conduit
auditif externe a une fréquence de
résonance de 2,5 kHz.
• La conque seule a une fréquence de
résonance vers 5,5 kHz.
• L’intensité d’un son sur le tympan est
multiplié par un facteur 10 dans cette
gamme de fréquence.

II) OREILLE MOYENNE
La membrane tympanique, zone de vibration impliquées :
• Aux basses fréquences, le déplacement de la membrane tympanique est grand et la transmission
augmentée.
• Aux plus hautes fréquences, la membrane tympanique "se sépare" en plusieurs zones moins efficaces
pour transmettre le son. Avec des hautes fréquences, l’amplitude de déplacement est grande mais ce
déplacement est « anarchique ».
La chaine ossiculaire :
• Les osselets assurent la transmssion de la vibration sonore à travers cavum tympani (milieu aérique)
vers l’oreille interne (milieu liquidien). Ils permettent l’amplification du signal mais aussi et surtout
suppriment cet interface acoustique. Le rapport entre deux impédances acoustiques (air/eau) est très
élevé : Z1/ Z2 = 1/ 3750.
• Mécanismes mis en jeu :
La pression est augmentée dans le rapport des surfaces utiles des osselets (15 à 30 fois).
La vitesse de propagation diminue à partir de l’étrier (divisée par 1,3).
Les variations d'incurvation des parois du cône tympanique entraîne une augmentation de la force
(par 2,6) et une diminution de la vitesse.
• Au total
une augmentation de pression d'environ 45 fois = un gain de 35 dB
Fonction de protection :
• La contraction réflexe des muscles de l'oreille moyenne atténue sélectivement les composantes de
basse fréquence (inférièures à 1000 - 2000 Hz) et permet donc une meilleure perception des stimuli
complexes aux intensités élevées ainsi d’une protection de l’oreille interne.
• Ce réflexe bilatéral, polysynaptique présente un seuil à 70 - 90 dB au dessus du seuil de l'audition,
diminuant chez les personnes âgées : il augmente pour les hautes fréquences, ou pour des stimuli
brefs.
• La contraction du stapédius est également provoquée par la vocalisation, la réponse du muscle
précédant l'émission vocale.
• Le tympan(4) sépare le conduit auditif
externe de la cavité de l'oreille moyenne qui
est en relation avec la cavité buccale par la
trompe d'Eustache (6).
• La fenêtre ovale, sur laquelle s'applique la
platine de l'étrier (3), et la fenêtre ronde (5)
séparent oreille moyenne et oreille interne.
• La chaîne ossiculaire comprend le marteau
(1), l'enclume (2) et l'étrier (3) : elle relie le
tympan à la fenêtre ovale.

III) OREILLE INTERNE
La cochlée (ou limaçon) se présente comme un tube tronconique (la columelle) enroulé autour d'un axe
creux, de 35 mm de long, le Modiolus, sur deux tours trois quart chez l'être humain.Sur une coupe
transversale, le tube cochléaire apparaît divisé en trois canaux : canal cochléaire contenant l'endolymphe,
celui des rampes vestibulaire et tympanique contenant la périlymphe. Au centre (modiolus) le ganglion
spiral et les fibres du nerf cochléaire.
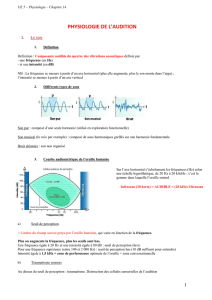
a) Tonotopie
Von Békesy (Prix Nobel en 1961) montra qu’une vibration de l'étrier provoque une vibration de la
membrane basilaire entraînée par la différence de pression hydraulique entre les deux rampes. En fonction
de sa fréquence, la vibration à un effet maximal (résonance) en un point différent de la membrane basilaire :
c'est la tonotopie passive.
Un son de fréquence élevée affecte une portion basale de la cochlée.
Un son de fréquence grave affecte une portion plus apicale de la cochlée (vers l’hélicotrema).
Tonotopie passive :
Puisque du fait de sa structure, la membrane basilaire est plus rigide à la base qu’au sommet, la déformation
est 10 000 fois plus importante au sommet. Les oscillations de pressions dans la rampe vestibulaire se
traduisent au niveau de la membrane basilaire par une onde propagée progressant de la base vers l’apex avec
une amplitude croissante. Puis, au delà d'un maximum, dont la position est fonction de la fréquence de la
stimulation, l'onde s'arrête brusquement. Ainsi plus la fréquence est basse, plus la déformation est grande
plus le nombre de cellules ciliées stimulées est grand et inversement.
Tonotopie cochléaire :
Rhodes montra le premier en 1978 que la réponse de la membrane basilaire n'était pas linéaire.
En effet, lorsque l'intensité d'un son (IdB) augmente, la valeur du rapport des déplacements de la membrane
basilaire sur celui du marteau n'est pas une constante.
Un grand nombre de chercheurs ont étudié le mode de vibration non pas de la membrane basilaire, mais du
complexe (Membrane basilaire + Organe de Corti), révélant des modes de vibration variant avec la
fréquence, et pour une fréquence avec l'intensité : c'est la tonotopie cochléaire.
• Le canal cochléaire (1) est isolé de la rampe
vestibulaire (2) et de la rampe tympanique (3).
• Il contient l'organe de Corti, dont les cellules
ciliées externes et internes vont assurer
l'amplification (CCE), la sélectivité, et en final
la transduction de l'énergie mécanique en
énergie électrique (CCI), c-à-d potentiel
d'action du nerf auditif.
• L'organe de Corti est recouvert par la
membrane tectoriale (6) flottant dans
l'endolymphe ; il repose sur la membrane
basilaire (5) au contact de la rampe tympanique
(3).
• La lame spirale osseuse (9) relie l'organe de
Corti au ganglion (8).

De toutes manières, ces 2 types de tonotopies sont passives, il n’y a pas de phénomènes actifs.
b) L’organe de Corti
Les cellules ciliées :
Les 4 rangées de cellules sont régulièrement espacées, tous les 10 µm, le long de la membrane basilaire.
Dans chaque rangée, on compte environ 100 cellules ciliées par mm, soit au total 3 500 cellules ciliées
internes et 12 000 cellules ciliées externes.
Il y a donc seulement 16 000 cellules ciliées dans la cochlée comparé aux 100 millions de photorécepteurs
dans l'œil.
De plus, les cellules ciliées ne se renouvellent pas.
L'organe de Corti est donc très fragile. Toutes les agressions se cumulent au cours du temps, pouvant
déclencher une surdité de perception.
Les cellules ciliées internes (CCI):
Les CCI,). Implantés au pôle apical de la plaque cuticulaire, une centaine de stéréocils sont alignés en 3 ou 4
rangées de taille croissante. Ils plongent en totalité dans l'endolymphe, MAIS ne sont pas en contact avec la
plaque tectoriale. La présence de filaments d'actine dans les stéréocils assure leur rigidité. A la base des CCI,
95 % des neurones du ganglion auditif font synapse. Les cellules ciliées internes sont les cellules assurant la
transduction mécano-électrique.
Les cellules ciliées externes (CCE):
Les CCE, cylindriques, 6 m de diamètre, ont une longueur passant de 25 m au tour basal (Perception des
Hautes Fréquences) à 70 m au tour apical (Perception des BF). Elles sont disposées en rangées parallèles
de la base au sommet de la cochlée ; le nombre de CCE par range varie selon les espèces de 3 à 5. Chaque
CCE est solidement ancrée sur la cellule de Deiters. Ce qui permet un couplage mécanique entre la
membrane basilaire et la membrane tectoriale.
• Dans cette section transversale de la partie
basale d'une cochlée de mammifère, 1 CCI (1)
et 3 CCEs (2) sont représentées de part et
d'autre du tunnel de Corti (3).
• La membrane tectoriale (6), flottant dans
l'endolymphe coiffe les stéréocils des cellules
ciliées.
• Les fibres nerveuses gagnent ou quittent
l'organe de Corti par l'habenula perforata (5) au
travers de la membrane basilaire (4).

Illustration du couplage mécanique :
c) Les cils
Implantés dans la plaque cuticulaire apicale ou lame réticulaire, les cils sont longs de 6 à 7 m et forment
un W pour les cellules ciliées externes.
Les stéréocils sont liés entre eux par des liens transversaux et apicaux dont l'intégrité fonctionnelle est
primordiale.
La flexion des stéréocils est à l'origine de la mécanotransduction.
Dans la membrane tectoriale, la flexion horizontale des cils vers l'extérieur, soit vers les plus grands, est à
l'origine de la dépolarisation cellulaire, tandis qu'une flexion vers les plus courts (vers l'intérieur), est à
l'origine de l'hyperpolarisation.
• Au niveau des CCI, la dépolarisation entraînera une augmentation de la décharge dans les fibres
afférentes du nerf auditif, proportionnelle à l'amplitude de la flexion.
• Au niveau des CCE, la dépolarisation entraînera un changement de longueur de la cellule,
proportionnelle à l’amplitude de la flexion.
Tonotopie active :
Les CCE sont donc capables à la fois de transmettre le mouvement de l'organe, et de produire des forces qui
agissent en retour sur cet organe selon un mode unique de mobilité cellulaire.
La contraction des CCE permet d’amplifier le signal à l’endroit où la vibration cochléaire (cf tonotopie
passive) est la plus importante. Ceci offre un gain de 50 Db.
• A la base, la CCE est courte, les stéréocils sont les plus longs et
fortement implantés dans la membrane tectoriale, la cellule de
Deiter est très rigide : un couplage ferme est donc réalisé entre
les membranes basilaire et tectoriale et la contraction de la
cellule ciliée externe peut modifier la mécanique cochléaire.
• A l'apex, la CCE est longue, simplement posée sur une cellule de
Deiter peu rigide et le couplage MB/MT est pratiquement
inexistant.
Les mouvements des liens apicaux constitués de myosine
permettent l'ouverture des canaux ioniques situés au sommet
des cils.
L'influx de K+ dans la cellule ciliée est responsable du
changement de potentiel membranaire, proportionnel à
l'intensité acoustique du son stimulant
 6
6
1
/
6
100%