Organisation et ajustement des commandes motrices.

Chapitre 1 : Organisation et ajustement des commandes motrices. 1
O
OR
RG
GA
AN
NI
IS
SA
AT
TI
IO
ON
N
E
ET
T
A
AJ
JU
US
ST
TE
EM
ME
EN
NT
T
D
DE
ES
S
C
CO
OM
MM
MA
AN
ND
DE
ES
S
M
MO
OT
TR
RI
IC
CE
ES
S
I. BASES NEUROPHYSIOLOGIQUES DU MOUVEMENT
A. LE MOUVEMENT
Pour tout mouvement, il y a 3 types d’activités motrices qui sont mises en
jeu simultanément.
1. ACTIVITES DYNAMIQUES
L’activité dynamique concerne directement le mouvement à réaliser
(frapper le ballon avec le pied, lancer une balle…)
Elles mobilisent globalement les articulations distales du corps (poignet,
cheville, genoux, coude)
Elles mobilisent au minimum 2 groupes musculaires. Un groupe que l’on dit
agonistes, qui permettent la rotation d’une articulation dans une direction de
l’espace et bien sûr pour stopper le mouvement il faudra mettre en jeu un groupe
de muscles antagonistes qui permettent de freiner le mouvement et de le stopper
là où c’est nécessaire.
Le système nerveux doit pouvoir gérer l’intensité d’activation de ces muscles et
le « timing » des activations.
2. LES REACTIONS D’EQUILIBRATION
Ces réactions ont pour objectif de maintenir la projection verticale du
centre de gravité dans le polygone de sustentation. Cela prend en compte les
forces de gravité qui sont détectées au niveau du système vestibulaire.
G G
Déstabilisation Ré-équilibration

Chapitre 1 : Organisation et ajustement des commandes motrices. 2
Rq : les mouvements respiratoires provoquent des réactions de ré-équilibration.
3. LES ACTIVITES POSTURALES
Ces activités posturales ont 2 objectifs :
- La stabilisation des segments corporels.
- L’orthostatisme, c’est à dire de maintenir une posture debout.
Les activités posturales concernent particulièrement les articulations proximales
du corps (tête, tronc, épaules, hanches)
Ajustement postural
G
Modification posturale liée au mouvement du bras.
Rq : Lorsqu’on lance un objet, afin d’éviter d’avoir une rotation du tronc qui suit
le mouvement et qui provoquera une déviation de la trajectoire, il y a une
adaptation nerveuse qui mettra en jeu des muscles de l’épaule et du tronc pour
ajuster la posture.
L’exécution d’un mouvement est un processus complexe qui nécessite la
coordination d’un ensemble d’activités musculaires au niveau d’un grand
nombre d’articulations qui ne sont pas forcement liées directement au
mouvement à réaliser.
B. ORGANISATION GENERALE DU CONTROLE MOTEUR
L’organisation centrale du mouvement se fait en 2 étapes :
- Planification et programmation du mouvement.
- Exécution du mouvement.
Cela concerne le transfert d’un signal de commande vers les moto-neurones des
muscles impliqués dans le mouvement mais aussi vers des structures qui sont
impliquées dans la régulation du mouvement afin de le corriger au cours de son
exécution si nécessaire.

Chapitre 1 : Organisation et ajustement des commandes motrices. 3
La planification du mouvement débute par la prise de décision qui va mettre en
route une série de traitement d’informations au niveau du système nerveux
central.
Ce traitement va permettre de préparer un programme moteur initial qui sera
envoyé vers les muscles concernés. En fait un programme moteur est
hiérarchisé, c’est à dire formé d’un ensemble de sous programmes eux-même
appelant d’autres sous-programmes plus simples.
PGM Moteur
général
PGM activité
membres
supérieurs
PGM activité
Tronc, tête
PGM activité
Membres
Inférieurs
PGM rotation épaule
PGM rotation coude
PGM rotation poignet
PGM rotation hanche
PGM rotation genou
PGM rotation cheville

Chapitre 1 : Organisation et ajustement des commandes motrices. 4
Pendant la réalisation du mouvement, le système nerveux central devra encore
continuellement traiter un ensemble d’informations pour savoir si le mouvement
réellement effectué est correct. S’il y a des divergences entre ce qui était
souhaité et ce qui est réalisé, il y aura une correction, un réajustement du
programme moteur. Ces ajustements permettront de contrôler avec précision la
durée, l’amplitude ou encore la vitesse d’un mouvement.
On emploie différents termes pour les ajustements : rétroactions, feedbacks,
réflexes, ajustement en boucles fermées.
Encéphale
Boucle supra-médullaire Signal de commande
Moelle épinière (moto-neurones)
Boucle médullaire
Muscle Système musculo-squelétique
Récepteur
Feedback Mouvement
Les ajustements du signal de commande peuvent se faire à deux niveaux. Au
niveau de la source, c’est à dire de l’encéphale grâce à ce qu’on appelle des
voies supra-médullaires ou la correction peut se faire à proximité des neurones
des muscles c’est à dire au niveau des circuits de la moelle épinière et on parle
de boucle médullaire.
Les ajustements supra-médullaires sont plus tardifs que les ajustements
médullaires. Par contre les ajustements médullaires sont nettement moins fins
que les ajustements médullaires. Ils vont générer des réactions relativement
brusques. C’est ce qui est à la source de mouvements saccadés.
Qui dit feedback, dit programme non optimal.

Chapitre 1 : Organisation et ajustement des commandes motrices. 5
Au cours d’un apprentissage moteur, on voit apparaître les ajustements
anticipatoires appelés feedforwards, ajustements en boucles ouvertes.
On les voit apparaître pour les ajustements de la posture nécessaire pour la
bonne réalisation d’un mouvement.
Au cours de la réalisation du geste, le système nerveux central apprend à
connaître la perturbation du mouvement et l’enregistre : automatisme du geste.
Programme postural Programme du mouvement
-
Feedback feedforward
Exécution de la posture Exécution du mouvement
Posture Mouvement
Perturbation
Ajustements posturaux tardifs
Ajustements anticipatoires
Tout ce qui est involontaire, ça peut être les feedforward et les feedback.
Tout ce qui est volontaire correspond à la prise de décision du mouvement et sa
mise en route.
Plus un geste est acquis, plus la part d’involontaire est importante.
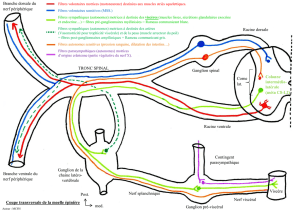
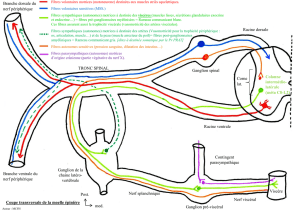
II. SUPPORT ANATOMO-FONCTIONNEL DU CONTROLE
MOTEUR
A. PRESENTATION GENERALE DU SYSTEME NERVEUX
CENTRAL, SCHEMAS SENSORI-MOTEURS
Le système nerveux est subdivisé en 2 grandes parties :
- L’encéphale.
- La moelle épinière.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
1
/
33
100%