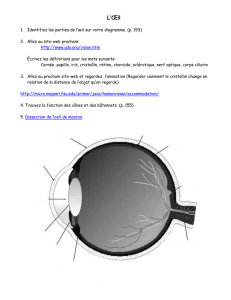
Formation de l`oeil

Analyse détaillée du phénomène d'induction
Formation de l'oeil
Comme il a été mentionné précédemment, les expériences d'embryologie expérimentale de
Spemann (transplantation de la lèvre du blastopore des amphibiens) ont mené à la découverte
du phénomène d'induction embryonnaire, et donc de l'importance de la communication entre
cellules et tissus au cours du développement. Les effets observés après cette transplantation
sont complexes puisqu’ils impliquent une cascade d’évènements s’enchaînant les uns derrière
les autres pour finalement aboutir à la formation d'un embryon ectopique complet. En fait,
Spemann avait analysé ce processus d'induction sur un modèle expérimental plus simple, la
formation du cristallin de l'oeil. Et c’est en observant ce processus developpemental qu’il
avait formalisé sa théorie de l’induction embryonnaire.
Description de la formation de l'oeil chez l'embryon
La formation du cristallin se fait à partir d'un épaississement (placode) de l'ectoderme de la
tête. Proximité de la vésicule optique (extension latérale du cerveau antérieur) et du cristallin.
Transformation de la vésicule optique en rétine neurale et rétine pigmentée.
A la fin du XIX è siècle, Spemann, ainsi que d'autres embryologistes avaient observé que chez
des embryons nés (naturellement) cyclopes, une rétine centrale était associée à un cristallin. La
formation d'une rétine ectopique était donc associée à la formation ectopique d'un cristallin.
L'idée a donc germé que la rétine serait responsable de l'induction du cristallin. Des expériences
de transplantations et extirpations de ces différents tissus ont ensuite pu démontrer que la
vésicule optique est en effet nécessaire à la formation du cristallin.

Ces mêmes expériences démontrent que la vésicule optique n'est pas suffisante pour induire la
formation d'un cristallin ectopique n'importe où dans l'embryon. Seul l'ectoderme de la tête
répond à l'induction de la vésicule optique. Cet ectoderme est donc compétent à répondre au
signal de la vésicule optique. Inversément, on voit que d'autres tissus que la vésicule optique ne
possèdent pas ce pouvoir d'induction. Donc notion de régionalisation de la compétence. On
peut analyser de manière expérimentale le phénomène d'induction du cristallin chez le Xénope.
Les résultats de ces expériences sont résumés sur le schéma ci-dessous.
A) Au moment de la gastrulation, tout l’ectoderme du pôle animal est compétent à se
transformer en cristallin, s’il est exposé à des signaux appropriés. L'expérience consiste à
découper la région dorsale de l'embryon au stade blastula (la calotte animale) et à l'exposer, en
culture, à divers inducteurs potentiels. En fait, cet ectoderme du pôle animal est successivement
compétent à se transformer en mésoderme, tissu neural, cristallin, quand il est exposé à des
signaux distincts. La période de compétence à se transformer en cristallin est courte (qques
heures). Ceci est également une règle du phénomène d'induction: il y a une fenêtre de temps
pendant laquelle celle-ci peut se produire.
B) Au stade neurula, cette induction n’est pas encore terminée; toutefois, on peut démontrer
qu’une large portion de l’ectoderme de la tête à une tendance à se transformer en cristallin
(toujours le test d'explants en culture), en ce sens que des signaux faibles parviennent à le
transformer en cristallin, alors qu’il faut des signaux forts pour transformer un ectoderme de
pôle animal en cristallin.
C) Au moment de la fermeture du tube neural, une portion restreinte de l’ectoderme de la tête
est spécifiée à se transformer en cristallin. Spécification, veut dire qu’on peut démontrer
expérimentalement que cet ectoderme en isolation (càd en culture) a la propriété de se
transformer en cristallin (quoique mal formé et petit) de manière autonome. A ce moment, les
protubérances latérales du cerveau antérieur (qui donneront les vésicules optiques) commencent
à peine à se rapprocher de l”ectoderme de la tête.

Plus tard seulement commence l’étape de croissance et différenciation du cristallin, qui continuera
pendant une bonne partie de la vie embryonnaire. On voit donc que la spécification du cristallin se
produit avant même que la vésicule optique n’ait rejoint l’ectoderme de la tête. Ainsi, la spécification
du cristallin semble en grande partie indépendante de la vésicule optique. Par contre, on peut
démontrer que la croissance et la différenciation du cristallin semblent entièrement dépendantes de
la présence de la vésicule optique.
Nous voyons ainsi de manière expérimentale que la formation du cristallin est un processus que l'on
peut décomposer en plusieurs étapes successives dont chacune a son importance dans la formation
des structures.
1) Une période où la compétence à former un cristallin se met en place dsans l'ectoderme de la
mid-gastrula.
2) L'acquisition par l'ectoderme d'une tendance / biais à former un cristallin au début de la
neurulation
3) La spécification de l'ectoderme à un destin "cristallin" en fin de neurulation
4) Une phase de différenciation et croissance du cristallin qui s'installe ensuite, pour perdurer
toute la vie.
Un niveau supplémentaire de complexité peut être observé au cours de la formation de l'oeil. C'est le
fait que le cristallin en formation (en fait la placode optique) est elle aussi la source d'un inducteur
agissant sur la vésicule optique elle-même. La placode est importante pour que la vésicule optique se
transforme en coupe à deux feuillets dont l'un est la rétine neurale, l'autre la rétine pigmentée. On voit
ainsi un phénomène d'induction réciproque de tissus l'un sur l'autre.
Conclusion sur le phénomène d'induction
Importance de l'aspect temporel et régional du phénomène d'induction (càd: i) la fenêtre de temps
pendant laquelle un tissu est compétent à répondre est réduite et ii) une région d'un tissu peut être
compétente à répondre, alors qu'une autre région ne l'est pas.
Un aspect primordial du phénomène d'induction est donc le contexte cellulaire dans lequel elle se
produit, qui correspond à "l'état" dans lequel un tissu est apte à répondre à un ou des signaux
particuliers.
Avantage d'un tel système: on peut utiliser les mêmes signaux à plusieurs reprises et avoir des
résultats différents, puisque le contexte cellulaire est différent.

Les molécules impliquées dans le phénomène d'induction
Plusieurs signaux ont été identifiés récemment qui jouent de multiples fonctions à différents moments
du développement.
Depuis plus de vingt ans que les chercheurs essayent de comprendre comment cet organe se forme,
plusieurs gènes jouant un rôle crucial au cours de son développement ont été identifiés. Parmi ceux-ci,
les facteurs de transcription Rx et Pax6. L'absence de Rx comme de Pax6 entraîne l'absence des yeux
chez les animaux. Ces gènes sont exprimés au cours du développement (expérience d'hybridation in
situ, qui met en évidence les cellules/tissus qui expriment l'ARN codant pour un gène) dans une
région antérieure continue de la plaque neurale et non dans deux champs distincts droite et gauche
correspondant aux deux yeux futurs.
Rx
Expression de Xrx dans la partie antérieure de la plaque neurale chez le
Xénope
KO de Rx chez la souris. Le KO de Rx chez la souris entraîne l'anophthalmie de ces
mutants (càd pas d'yeux). Chez ces mutants, la vésicule optique ne se forme pas.

Chez les humains, l'absence de Rx conduit à la même anomalie, que l'on appella anophthalmie.
L'anophtalmie est le défaut des yeux le plus fréquents chez l'homme et la cause principale de la cécité.
Pax6
Expression de Xpax6 dans la partie antérieure de la plaque neurale
chez le Xénope
Mutant rat pour le gène Pax6: pas d'yeux, mais également des malformations importantes
de la face (montrant que Pax6 a des rôles multiples au cours du développement de la tête.
Pax6 a été énormément étudié depuis sa découverte il y a dix ans. On a pu montrer que Pax6 est
exprimé non seulement aux étapes précoces du développement (comme montré chez le Xénope, mais
également plus tard. Par exemple, il est exprimé par la placode de l'oeil et par la vésicule optique. On
a pu montrer par des expériences de recombinaison de tissus (recombiner des tissus mutants avec des
tissus WT) que l'expression dans la placode est essentielle à la formation du cristallin.
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%