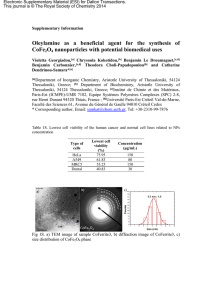
Asym 0

Divisions asymétriques chez les Angiospermes
De la première division du zygote, jusqu’à la mise en place des gamètes mâles et femelles,
des divisions asymétriques surviennent et permettent la mise en place de types cellulaires
différents, que ce soit dans le sporophyte ou dans le gamétophyte. Les exemples de divisions
asymétriques chez Arabidopsis thaliana débutent lors de l’embryogenèse précoce. En effet, et
bien que cela ne soit pas la règle pour toutes les Angiospermes (Kaplan et Cooke, 1997), la
première division du zygote est asymétrique en terme de taille et de devenir, puisque la cellule
apicale constituera la majeure partie de l’embryon mature, alors que la grande cellule basale
donnera le suspenseur dont seule la cellule la plus apicale participera à la construction d’une
partie de la racine. Durant le développement embryonnaire, d’autres divisions asymétriques
surviennent, notamment la division tangentielle des cellules apicales qui isole l’épiderme
(protoderme alors) des cellules sous-jacentes, au stade 8-12 cellules. De même, un peu plus
tard, une division asymétrique met en place dans l’hypophyse (provenant de la cellule la plus
apicale du suspenseur), la cellule lenticulaire, qui est la cellule mère du centre quiescent.
L’initiation des racines secondaires à partir de deux cellules « superposées » du péricycle
requiert leur division asymétrique , produisant deux petites cellules adjacentes et deux grandes
cellules éloignées (Casimiro et al., 2003). Dans ce cas, ce sont des cellules qui avaient cessé
d’être cyclantes qui sont stimulées pour entre à nouveau dans le cycle cellulaire.
Le méristème racinaire possède des cellules initiales (cellules souches) qui se divisent de
façon asymétrique pour mettre en place deux cellules sœurs, l’une conservant la qualité de
cellule initiale, l’autre va se différencier pour mettre en place une assise cellulaire. Dans le cas
de l’initiale des cellules du cortex et de l’endoderme, une cellule souche se divise une
première fois, produisant deux cellules sœurs, l’une demeurant initiale, l’autre va se cliver à
nouveau, perpendiculairement à l’axe de la première division, donnant ainsi deux cellules
différentes en taille, une plus fine, vers le centre, à l’origine de l’endoderme, l’autre plus
épaisse, vers l’extérieur qui donnera le cortex (cf figure).
En ce qui concerne la gamétogenèse, du côté mâle, la division de la microspore est
asymétrique et donne deux cellules de tailles différentes, l’une complètement englobée dans
l’autre, avec des noyaux de tailles différentes. Lors de la mise en place du sac embryonnaire,
une très grande cellule est produite, la cellule centrale.

Dans la feuille, les stomates sont mis en place à la fin d’un processus qui débute par une
division asymétrique isolant le méristémoïde (un méristème secondaire ?) des cellules en
puzzle (Larkin et al., 1997).
Ce n’est que très récemment que Wang et al. (2007) ont montré le rôle d’un module MAP
kinase dans le contrôle de cette division asymétrique et de la première division (asymétrique)
du zygote. D’autres méristème secondaires possèdent des cellules qui se divisent de façon
asymétrique. Chez un arbre vivant parfois quelques milliers d’années, la croissance en
épaisseur (parfois plusieurs mètres) est assurée par le fonctionnement du cambium. Cette
assise produit le bois secondaire vers le centre et le liber secondaire vers l’extérieur (donc des
cellules à devenirs différents). Enfin, des formations secondaires péridermiques (comme le
liège ou suber) résultent du fonctionnement d’une assise génératrice, le phellogène (ou plus
pompeusement assise subéro-phellodermique) qui produit le liège vers l’extérieur et le
phelloderme vers l’intérieur, là encore des tissus différents.
Bergmann DC, Lukowitz W, Somerville CR 2004 Stomatal development and pattern
controlled by a MAPKK kinase. Science 304: 1494-1497
Casimiro I, Beeckman T, Graham N, Bhalerao R, Zhang H, Casero P, Sandberg G, Bennett
MJ 2003 Dissecting Arabidopsis lateral root development. Trends Plant Sci 8: 165-171
Kaplan DR, Cooke TJ 1997 Fundamental Concepts in the Embryogenesis of
Dicotyledons: A Morphological Interpretation of Embryo Mutants. Plant Cell 9:
1903-1919
Larkin JC, Marks MD, Nadeau J, Sack F 1997 Epidermal cell fate and patterning in leaves.
The Plant Cell 9: 1109-1120
Lukowitz W, Roeder A, Parmenter D, Somerville C 2004 A MAPKK kinase gene
regulates extra-embryonic cell fate in Arabidopsis. Cell 116: 109-119
MacAlister CA, Ohashi-Ito K, Bergmann DC 2007 Transcription factor control of
asymmetric cell divisions that establish the stomatal lineage. Nature 445: 537-540
Pillitteri LJ, Sloan DB, Bogenschutz NL, Torii KU 2007 Termination of asymmetric cell
division and differentiation of stomata. Nature 445: 501-505
van den Berg C, Willemsen V, Hage W, Weisbeek P, Scheres B 1995 Cell fate in the
Arabidopsis root meristem determined by directional signalling. Nature 378: 62-65
Wang H, Ngwenyama N, Liu Y, Walker JC, Zhang S2007 Stomatal development and
patterning are regulated by environmentally responsive mitogen-activated protein
kinases in Arabidopsis. Plant Cell 19: 63-73
Xiao W, Custard KD, Brown RC, Lemmon BE, Harada JJ, Goldberg RB, Fischer RL
2006 DNA methylation is critical for Arabidopsis embryogenesis and seed viability.
Plant Cell 18: 805-814
1
/
2
100%